
THE PENYCUIK EXPERIMENTS.
CONTENTS.
GENERAL INTRODUCTION
(1) Zebra Hybrids. (2) The Principles of Breeding - (a) Reversion in Pigeons, Fowls, Dogs, Rabbits, Ducks, and Horses; (b) Prepotency and Inbreeding. (3) Telegony. (4) Saturation. (5) Sterility in Equine Hybrids.
PART I. A. The Birth of a Hybrid between a Burchell's Zebra and a Mare (This paper is reprinted with a few verbal alterations from The Veterinarian for November, 1896.)
B. Zebra- horse Hybrids . (This paper is reprinted from The Zoologist for February, 1898.)
PART II. Telegony and Reversion(This paper in an abridged form was published in The Veterinarian, November and December, 1897, and February, 1898.)
APPENDIX. Telegony (Reprinted from The Veterinarian, April and May, 1895.)
LIST OF ILLUSTRATIONS.
1. Matopo, a Burchell zebra
2. Mulatto, a West Highland pony
3. Mulatto and her hybrid Romulus seven days old
4. Romulus, twenty-seven days old
5. Romulus, twenty-seven days old, face view
6 & 7. Matopo, left side
8. Matopo, right side
9. Mulatto, and Romulus seven days old
10. Romulus, view of face
11. Romulus, one year old
12. Brenda, a Clydesdale mare's hybrid
13. Lord Morton's Quagga
14. Lord Morton's Quagga hybrid
15. Filly out of Lord Morton's mare .
16. Mulatto
17. Matopo, left side
18. Matopo, right side
19. Matopo, face markings
20. Romulus, face markings
21. A Somali zebra
22. Skin of a Somali zebra, from a photograph of a skin lent by Mr. Charles V. A. Peel, Oxford
23. Diagram of stripes of Matopo
24. Diagram of stripes of a Somali zebra
25. A zebra filly with "shadow" stripes, from a photograph by Mr. Brown, Penycuik
26. A mountain zebra
27. A true Burchell zebra
28. Skin of a mountain zebra
29. Skin of a Crawshay's zebra
30. Mountain zebra and foal
31. Mulatto, and Romulus seven days old
32. Romulus twenty-seven days old
33. Romulus twenty-seven days old, face view
34. Somali zebra, face markings
35. Crawshay's zebra, face markings
36. Face markings in a Norwegian pony
37. A zebra dam, horse sire, hybrid
38. Skin of dam of hybrid shown in Fig. 37
39. Sire of hybrid shown in Fig. 37
40. Mulatto
41. A grey Arab horse (Benazrek)
42. An ordinary mule - ass sire, horse dam - hybrid
43. Mulatto's second foal, from a drawing by Mr. Edwin Alexander
44 Mulatto's second foal
45. Ridges formed by hair in region of flank feather in a foal
46. Part of skin shown in Fig. 45
Figs. 1, 4, 5, and 42, from photographs by Mr. Charles Reid, Wishaw. Figs. 3, 6, 11, 12, and 41, from photographs by Mr. Swan Watson, Edinburgh . Figs. 2 and 44, from photographs by Mr. Addcrley, Penycuik. Figs. 19, 20, 23, 24, 36, 45, and 46, from drawings by Mr. Fiddes Watt. Figs. 13, 14, and 15.- 13 by Mr. Fiddes Watt; 14 and 15 by Mr. R. E. Holding, after Agasse's pictures in Roy. Coll. of Surg. Mus. London. Figs. 21, 26, 27, 34, and 35, from drawings by Mr. J. Smit, in Prazak's ' Wild Horses of Africa,' MS. Figs. 28, 29, 37, 38, and 39, from photographs by Mr. Fall, Baker Street, London. Fig. 30, from a photograph kindly sent by the Director of the Zoological Gardens, Amsterdam. The foal is now in the Zoological Gardens, London. For the use of the skins represented in Figs 28 and 29 I am indebted to Mr. Rowland Ward, Piccadilly, London.
2024 PREFACE
A COMMUNICATION OF A SINGULAR FACT IN NATURAL HISTORY. BY THE RIGHT HONOURABLE THE EARL OF MORTON, F. R. S. IN A LETTER ADDRESSED TO THE PRESIDENT
Philosophical Transactions of the Royal Society, 31 December 1821, Volume 111
My Dear Sir, I yesterday had an opportunity of observing a singular fact in Natural History, which you may perhaps deem not unworthy of being communicated to the Royal Society, Some years ago, I was desirous of trying the experiment of domesticating the Quagga, and endeavoured to procure some individuals of that species. I obtained a male; but being disappointed of a female, I tried to breed from the male quagga and a young chesnut mare of seven-eighths Arabian blood, and which had never been bred from: the result was the production of a female hybrid, now five years old, and bearing, both in her form and in her colour, very decided indications of her mixed origin. I subsequently parted with the seven-eighths Arabian mare to Sir Gore Ouseley, who has bred from her by a very fine black Arabian horse. I yesterday morning examined the produce, namely, a two-years old filly, and a year-old colt. They have the character of the Arabian breed as decidedly as can be expected, where fifteen-sixteenths of the blood are Arabian; and they are fine specimens of that breed; but both in their colour, and in the hair of their manes, they have a striking resemblance to the quagga. Their colour is bay, marked more or less like the quagga in a darker tint. Both are distinguished by the dark line along the ridge of the back, the dark stripes across the fore-hand, and the dark bars across the back part of the legs. The stripes across the fore-hand of the colt are confined to the withers, and to the part of the neck next to them; those on the filly cover nearly the whole of the neck and the back, as far as the flanks. The colour of her coat on the neck adjoining to the mane is pale, and approaching to dun, rendering the stripes there more conspicuous than those on the colt. The same pale tint appears in a less degree on the rump; and in this circumstance of the dun tint also she resembles the quagga.

GENERAL INTRODUCTION.
I HAVE been frequently asked when an account is likely to be published of the telegony and other experiments I started some time ago at Penycuik.* As the problems under consideration are not of a kind that can be settled off-haudj and as one inquiry has begotten others, some years must elapse before a complete and systematic account is possible. [* Penycuik is pronounced (as it was often formerly spelt) Pennycook. It means the hill of the cuckoo. Now the usual spelling is Penicuik, but until comparatively recently, as in olden times, a "y" was almost invariably used. Whether the Post Office or the North British Railway Company deserves most blame for the change it is hard to tell.]
For the information of those good enough to interest themselves in some of the problems I have set myself to solve, I have decided to issue in book form three papers which have already been published. These papers, with the help of this introduction (which must also serve as a preface, and to a certain extent as a supplement), will indicate the lines along which the inquiries are proceeding, and also the kind of answers likely eventually to be made to some of the questions. The first two papers deal with five of the zebra hybrids I have bred; the third - which has grown out of an address prepared nearly a year ago - deals chiefly with reversion and telegony.
The three papers having been written at different times and for different objects, a certain amount of repetition has been inevitable. This will, I think, prove advantageous rather than otherwise; it will at least help in fixing in the mind of the reader the difference in the arrangement of the stripes in the Burchell zebra stallion "Matopo" from that in his hybrid offspring.
When I came to seriously consider the "infection" or telegony problem, it was at once evident that a special study of the bands, stripes, and other markings in the Equidae was indispensable. Though much has been written bearing on the colouration of zebras, I failed to find anything- approaching a full account of the plan of the stripes in even the mountain zebra (Equus zebra), which has been almost constantly under observation in England for more than a century. Hence the necessity of working up, as far as the available material permitted, the whole colouration question in the Equidae, and hence it is that so much apace is occupied with a description of the bands and stripes in my zebra stallion, and with pointing- out how he differs from other zebras and from zebra hybrids.
It will be gathered from the first paper that I started with the idea that there are three kinds of zebras, each having its own distinctive characters. I still believe that there are three distinct types of zebras, but I no longer believe that all three types can be readily distinguished from each other by their markings. The three types are
(1) Grevy's zebra [E. grevyi) of Shoa and Somaliland [* It was doubtless the Shoa zebra, and not, as has hitherto been taken for granted, the mountain zebra of South Africa, that was exhibited in the Roman amphitheatre during the third century of the present era.]
(2) the mountain zebra [E. zehra), once common in South Africa, and generally referred to in old works as "the zebra," or "the common zebra;" and (3) the widely distributed Burchell group of zebras, including many species - or, at least, sub-species and varieties, - some of which are unfortunately often spoken of as quaggas. At the risk of iteration I may add that the Somali or Grevy's zebra (for a time confounded with the mountain zebra of South Africa) stands apart from all the others. It is profusely striped, measures sometimes quite fifteen hands (sixty inches), and is, I think, the most primitive of all the living striped horses. I considered the mountain zebra equally distinctive until some months ago, when I came across a skin which seems to connect this long-known species with certain less familiar members of the great Burchell group found in Nyassaland. There is hence no link now wanting in the chain (if the striping alone is considered) that has at one end the common mountain zebra with a "gridiron" over the hind quarters and legs barred to the hoofs, and the true Burchell zebra, with perfectly white legs and scarcely a vestige of transverse stripes across either croup or loins. From the true Burchell zebra it is but a step to the better marked specimens of the now lost if not extinct quagga (E. quagga). How the members of the Burchelli group will be eventually arranged does not concern us, as we are fortunately no longer expected to believe in the fixity of species.
Quite as much attention might have been directed to the form, &c., of the hair as to the arrangement of the stripes in the Equidae. But an exhaustive study of the form, colour, and length of the hair of the Equidse and their hybrids at different seasons and in different areas would occupy a lifetime." [* As it happens, Nathusius has already done admirable work in this field, and I trust he may yet increase our obligations by new observations and discoveries. Nathusius finds that in its hair the Somali zebra stands apart from all the rest, and that in zebra hybrids the hair neither resembles that of their parents, nor yet does it occupy an intermediate position. ]
ZEBRA HYBRIDS.
I have bred nine zebra hybrids by crossing mares of various sizes (from 11 to 15 hands) and breeds with my zebra stallion "Matopo," and I have in my possession three hybrids out of zebra mares, one having for his sire a donkey, the other two were sired by ponies. The hybrids are to me especially interesting because of the curious blending of characters, derived apparently partly from their actual and partly from their remote ancestors, and because they shed new light on many questions of general interest, such as the origin of stripes, reversion, interbreeding, and prepotency. The reversion hypothesis or dogma is an extremely fascinating one. It has long interested breeders of all kinds, yet it is still shrouded in mystery. It is commonly believed that a child sometimes, instead of taking after its father, closely resembles its father's mother, or is the image of its own mother; but we are still a long way from accounting for such phenomena. As is pointed out in the second paper, some of the hybrids in make and disposition strongly suggest their zebra sire, others their respective dams; but even the most zebra-like in form are utterly unlike their sire in their markings. It is not a matter of taking after a grandparent, but after an ancestor in all probability thousands of generations removed, an ancestor probably far more like the Somali than any of the Bur- chell zebras.

Closely connected with reversion is the question of prepotency. It is generally assumed that an old species or variety is prepotent over a more recent species or variety. It is impossible to say whether zebra hybrids in their markings take after a remote zebra ancestor, or after an ancestor common to both zebras and horses, or after a hypothetical mid-parent combining the characters of the less remote ancestors of both zebras and horses. There is, however, no difficulty in seeing that while some zebra hybrids, apart from their stripes, closely resemble the zebra parent, others take after their horse parent, thus showing that the wild sire is not necessarily the most prepotent. But even when the hybrids are distinctly horse-like they never repeat recently acquired peculiarities, such as a blaze or short ears, high withers, or a small head and long neck.
It would not, e.g., judging from the results already obtained, be possible to breed from a piebald mare a hybrid showing patches of white; even from a mare that produced a piebald foal to a whole- coloured horse. This, I think, is not because of the prepotency of the wild sire, but because there is reversion at least in the body colour towards the wild and never gaudily coloured ancestors of the horse, as well as to the primitively decorated ancestors of the zebra.
In addition to throwing light on reversion and prepotency, the hybrids are interesting in being, in some respects, almost intermediate between their parents. In Romulus the mane is nearly intermediate; in Remus it is always upright; while in two others it is long and almost horse-like in winter, but short and upright in summer. Again, the warts (chestnuts) are sometimes intermediate. In some they are quite absent, as in zebras, from the hind legs; in others they are present, but less than half the usual size, or there is only one present, less than a third the usual size. The warts on the fore-legs may be large, oval, and smooth, as in the zebra sire; or smaller, rounded in form, and half as prominent as in the horse. In the same way the hoofs vary, being almost intermediate in some, zebra-like in others.
Romulus, the oldest of my hybrids, measured at the withers on his second birthday nearly as much as his sire. In many respects he resembles his dam (Mulatto), the black Highland pony; in some respects he is a zebra. In the head and neck he is less like Mulatto than ordinary Highland ponies, as if he took after some of Mulatto's less refined Highland ancestors. From the shoulders backwards the hybrid is getting more and more like his dam, and less like his zebra sire, in which the sides are flattened, while the muscles of the back stand out at each side of a median spinal g-roove. Mounted on perfect legs, Romulus moves freely and rapidly, and often carries himself as proudly as a zebra. He has already been in harness, and, being very docile, he ought when older to prove as useful as he is attractive.
While in make the horse predominates in Romulus, he takes after the zebra in his constitution and habits. A zebra is in all respects more intense than a horse, he is more on the alert, more timid and suspicious, and yet more imbued with curiosity. When he once decides to take action he moves more rapidly than a horse, is more regardless of consequences, and usually suffers less from an injury than from the shock to his nervous system. One evening, when Matopo had refused for hours to retire into his box, I threw a rug over him. He bounded across the court (first nearly upsetting a mare, and then nearly breaking his neck against the fence), and then rushed around his box until the dreaded object was got rid of. Had it been a lion the panic could hardly have been greater, but he soon learned that a rug, however suggestive, was perfectly harmless. Strangely enough, when a zebra-skin is hung up within sight of Matopo he becomes excited. He arches his neck as if preparing for single combat, and, though he refuses to approach the skin, he is evidently not consumed with fear, as is the case when a coil of rope is carried up to him. It has been suggested that the rope appeals to his instinctive dread of serpents, but I consider this unlikely. A serpent-like object he strikes at with his hoofs; when his legs are touched with a rope he drops on his knees or lies down altogether.
The other day Matopo escaped from his paddock, and eventually found his way down three grass steps on to a lawn. On two sides the lawn is bounded by a fairly steep shrub-clad bank, on the other two it slopes somewhat rapidly to a footpath. Nothing would induce Matopo to descend this slope. He galloped gaily round the lawn as if he were in a circus, and was with difficulty persuaded to return the way he came. A somewhat timid Exmoor pony went up and down the slope, on to and off the lawn, without any hesitation. Probably the zebra, had he been left to himself long enough, would have made a complete inspection of the garden.
The behaviour of the zebra interested me, because his hybrid son Romulus had previously conducted himself in a similar fashion when taken on to the lawn to be photographed. Romulus eventually, tempted by corn, allowed himself to be led away from the lawn along a narrow grass border from which return, without leaving the grass, was impossible. Though Romulus in many ways behaves like his sire, he is, compared with most zebras, "as water unto wine;" and, though it may take longer to break zebra hybrids than horses, they will, I believe, be more amenable than ordinary mules, and, if one may judge from a single case, infinitely more easily managed than zebra-ass hybrids. I may here mention that the figures illustrating the papers give a very imperfect idea of Romulus and the other hybrids. They are generally considered more attractive than their respective dams, and handsomer than their gaily painted wild sire.
Remus, the bay Irish mare's hybrid, is, though only a yearling, already taller than Romulus. Notwithstanding the longish head and upright mane, he is in many ways very like his dam. The body colour is bay; the stripes, which are abundant and pronounced, and, except on the forehead, arranged as in Romulus, are of a dark brown colour. Before completing his first year he had formed for himself a small herd of fillies, which he guards jealously from all intruders. As a foal his curiosity was so great that if I knelt down and looked at a particular tuft of grass he came up, and with lowered head gazed steadfastly in the same direction.
Brenda, though out of a larger mare, is smaller than Remus, more heavily built, and much lighter in colour. With the exception of her unusually long mule-like ears and low withers, she is more a horse than a zebra, in gait and disposition as well as in form. Heckla, the offspring of a yellow and white Iceland pony, has from the first been dark in colour, and but faintly striped on the neck and over the hind quarters. Apart from her stripes, mule-like ears and tail, she is not unlike a pony. Strangely enough she steps high like a hackney, though her dam almost invariably moves like a pacer. Norette, the offspring of a small black Shetland pony, is extremely zebra-like throughout. In the form of the head, neck, and hind quarters she is remarkably like a typical Burchell's zebra I saw recently, and she sometimes reminds me forcibly of some of the photographs of the lost quagga. During the winter Norette's mane was long and hanging to both sides, as well as freely over her brow, but during the summer it has been as short as in her zebra sire. Like her dam she had a very thick winter coat, consisting of long hairs; but her summer coat is smooth and shining, and made up of quite short hairs. Though very like a zebra in make, this hybrid moves in a somewhat different fashion from her sire. This may be partly owing to her hoofs being unusually long, - they are relatively longer and more pointed than in Matopo.
During the present summer four new hybrids by Matopo made their appearance. Brenda's dam produced a second hybrid which has led to no little speculation. Brenda, but faintly striped, might, at a little distance, were it not for her long ears and low withers, be taken for a half-bred yearling colt. The new hybrid (Brenda's own sister) is very dark in colour and quite a zebra in action and habits. Last year some accounted for Brenda showing little of the zebra by saying her dam, the small cross-bred Clydesdale, was inbred, and they now account for her full sister (Black Agnes) being zebra-like by saying their common dam has been "infected," or, to use Bruce Lowe's term, "saturated," by their common sire Matopo. Should the Clydesdale mare have a third hybrid, and then an ordinary foal, something definite may eventually be established, both as to "saturation" and telegony.
But Black Agnes is interesting for yet another reason. In Mulatto's second foal, as is fully described in the telegony paper, there were several stripes across the croup, at nearly right angles to the spinal ridge. In foals of mares that have neither had offspring to zebras nor asses, markings are at times seen across the hind quarters - subtle bands, not due, as a rule, to colour, but rather like water-marks on silk. Again, in one of Lady Meux's hybrids out of a zebra mare, and in two dun-coloured cobs I have had under observation, there are vestiges of fairly broad stripes across the loins and croup - stripes more like the bars of the gridiron in the mountain zebra than the narrow zigzag lines in Romulus and some of my other hybrids. In Black Agnes there were, at birth, "water-marks" and faint stripes over the hind quarters as well as the characteristic hybrid stripes and rows of spots. We had, as it were, a double photograph showing faintly the markings which seem to belong to the ancestors of the true horses, overlying stripes and spots presumably- inherited from the ancestors of the zebras. In other words, in the markings o£ Black Agnes we seem to have a reversion towards the ancestors of the dam as well as the sire.* Whether any vestiges of the double printing will be visible next spring it is impossible to say. [* In zebra-ass hybrids the spinal and shoulder stripes and the leg bars are evidently inherited through the donkey sire, not, as might have been expected, from the zebra dam.]
An unexpected event this summer was the birth of twin hybrids - a colt and filly. Unfortunately one of the twins, the filly, though well developed and perfect in every way, died almost as soon as it was born. The dam of the twins is a 14.2 hands thoroughbred, light chestnut mare, with high withers and a prominent breast-bone. The surviving twin, though striped like Romulus, resembles his dam in the body colour, but he will probably become darker as he gets older. He already, though in many ways like a zebra, looks quite different from his half-sister Black Agnes. But, however much they differ in appearance, they closely resemble each other in their attitudes and movements. Like a hinny foal (a hybrid between an ass and a Welsh pony stallion) they step high when trotting, and if any one is near, the head is carried nearly on a level with the neck but to one side, apparently that they may keep an eye on the intruder. At the gallop the head is carried in the middle line, but as soon as they break into a trot the head is moved from one side to another by way of determining the position of the possible enemy. It has been said the gallop is the natural pace of the horse. This I very much doubt; it is certainly not the ordinary pace of a zebra. When pursued by enemies, and when advancing to attack a rival, zebras move at the gallop, but under ordinary circumstances they proceed at a leisurely dignified trot.
Some of my hybrids in their movements are not unlike their sire, but the two foals just referred to remind me more of a young stag I watched trotting round a paddock during the present summer. Whether this very elegant action will be maintained as they increase in size remains to be seen.
I may here mention the surviving twin has helped to assure me that hybrids may have as marvellous powers of recovering from severe injuries as zebras. Two years ago a zebra mare dragged from its place a heavy iron feeding-trough. The trough was soon broken, and as the mare reared and swung the trough about her loose box, ere the rope could be severed her fore-limbs were cut and bruised to an alarming extent. In a few days she was all right again. The wounds healed without suppurating, and without any swelling of the limbs. Quite recently Matopo, when turning rapidly on the way from his paddock, came into violent contact with one of the rails of an upright fence. The skin between the two halves of the lower jaw was torn so as to form a deep pocket large enough to hold a walnut. Stitching was out of the question, but in a few days the pocket was closed, and the wound has mended so well that no scar or irregularity in the striping can be detected.
The twin was found one morning, when about two months old, with a flap of skin five inches long, and averaging one and a half inches in width, hanging down over the front of the left fetlock. The skin was replaced and stitched along one side, but the upper part being dead was cut off some days later. For a time the wound looked hopeless enough, but now it is quite healed, and only a very small scar is left. As in the zebra mare, there has been no lameness, and from first to last a complete absence of swelling either below the wound at the fetlock or above in the vicinity of the knee. [* During the stitching the little hybrid fought desperately, and cried piteously, sometimes uttering short barking sounds or crying like a wounded hare.]

One of this summer's hybrids unfortunately died when hardly three months old. The dam in this case was a young bay, small thoroughbred Irish mare. Mules are said to be hard to rear, but so far, with the exception mentioned, there has been no difficulty with my hybrids. The hybrid that succumbed was very faintly marked, and in most respects more a horse than a zebra. The dam, barely three years old when the hybrid was born, suffered from the Strongylus scourge during the winter, and was consequently out of condition during the spring. Moreover I believe the dam is a victim of inbreeding. One of the evils of inbreeding is that the young are often difficult to rear, succumbing sometimes to one disease, sometimes to another. This hybrid, though weak to start with, did extremely well for a time, but after a few days of unfavourable weather it took ill and died quite suddenly from what seemed to be an attack of acute rheumatism. Several other foals, having quite as well bred dams, living under exactly similar conditions, suffered not at all.
Before leaving the hybrids, I may say that since Romulus appeared on the scene Baron de Parana has succeeded in crossing a Burchell zebra and an ordinary mare in Brazil. Baron de Parana seems to have satisfied himself that all attempts to cross a male Burchell zebra and a mare prior to 1892 (when he began his experiments) had failed. Notwithstanding this, he set to work and eventually succeeded, his hybrid arriving not many months after the birth of Romulus. Though summer succeeded summer without the expected results, the Baron, with exemplary patience, continued his experiments until success crowned his efforts. All who have had experience in work of this kind will heartily congratulate Baron de Parana, and wish him still further success.
THE PRINCIPLES OF BREEDING.
The uncertainties of stock-breeding are proverbial, and fanciers are seldom surprised when the unexpected happens. We speak of the principles of breeding and of scientific breeders, but it looks often as if there were no principles, and as if in breeding the scientific method is inapplicable. Why of all vocations should the breeder's be the most uncertain? or shall the question rather be, why of all men should breeders be the most unreasonable, more than all others desirous to "eat their cake and have it"? The fundamental reason doubtless is that wherever there is life there is variation in one or more directions. Variation is welcomed by breeders and fanciers up to a certain point, but beyond this it is considered a positive disadvantage. When a desired strain, variety, or breed has been formed the breeder would like to arrest all further variation. As with a key an electric current is switched off, he would like to arrest further change. But not only has variation to be contended with, there is reversion and the evils that flow from inbreeding. Even when making a new strain much patient work may be lost through prepotency or sterility unexpectedly supervening where least wanted.
Can science do anything to make the work of the breeder less uncertain and haphazard? Can it help in the production of new forms, and stereotype them when once realised, so that they may be reproduced, "repeated" with as much certainty as statues are turned out of a mould? Science is incapable of either creating or, except to a limited extent, controlling life, but yet it may help the breeder to so influence the vital forces in operation that the desired goal is reached without un- necessary waste of time or energy. To all who give a moment's thought to the subject it will be evident that variation, reversion, inbreeding, and prepotency have to be especially reckoned with. Science, in as far as it throws light on these subjects, will doubtless be useful. On the great and all-important subject of "variation" my experiments have no direct bearing, but they justify my referring shortly to reversion, prepotency, and inbreeding.
1. Reversion. - Nearly ten years have elapsed since Mr. Galton's work on 'Natural Inheritance' appeared, and yet but few breeders seem to fully realise that the offspring often take not so much after their immediate as after their less remote ancestors, that probably in all cases there is more or less marked "regression towards mediocrity." If there is any truth in evolution, it is extremely probable that the protoplasmic units of which any given individual is composed have to encounter and overcome during development countless numbers of corresponding units representing the component parts of a long series of lost ancestors. If for any reason units representing the latest development of the race or variety - the latest variations, mental or physical - are unable to assert themselves, are lacking in power or stability, older and more potent units will take their place, and thus lead to regression or reversion. The reversion may be to recent, remote, or intermediate ancestors, and the tendency will in most cases be to revert to "sports" that here and there mark the route along which the development has proceeded.
Any two individuals my be likened to two pictures. Even if portraits by the same artist of two members of the same family, they would differ in form as well as in colour. Were it possible to blend two such pictures into one, the result would be a picture differing from the originals, but not necessarily intermediate between them. Some of the pigments (having been differently made up) might neutralise each other, and latent colours - colours which, for some reason or other, had been obscured in one or both of the originals - might come to the surface. There would be a loss of the characters which served to distinguish the individuals from each other and the rest of their relations, but yet there would doubtless be hints here and there of the originals, and, because of the blending, suggestions of some of their ancestors. The offspring of two individuals (unless very closely related) differ in much the same way, only the possibilities for variation are infinitely greater. The raw material, the living clay or protoplasm, out of which the new individual is developed, comes, as it were, from different quarries, with in each case a different history. In the struggle amongst the parts, as the development proceeds the legionaries of the immediate ancestors are sometimes worsted by those of the less recent ancestors, the grandparents being victorious in the centre, while still more remote ancestors secure occasional successes in the wings. The final result is in most cases a drawn battle, neither particularly satisfactory to those immediately concerned, nor to the world at large.
According to Mr. Galton's law of ancestral heredity, the two parents contribute half, the four grandparents one fourth, the eight great-grandparents one eighth, and so on, of the total heritage of the average offspring. It is, however, conceivable that, owing to what, for want of a better name, may be called antagonism between the protoplasmic units during fertilisation and development, the grandparents, or even the great-grandparents, might contribute more than the immediate parents, and that, when the "antagonism" is still more pronounced, the comparatively remote ancestors might become the main contributors. Darwin was the first to point out that the offspring of parents belonging to different species, or distinct varieties, do not, as might have been expected, occupy an intermediate position, but take after a remote ancestor. Now-a-days the possibility of reversion is sometimes called in question. I have nevertheless endeavoured to account for my hybrids differing in their markings from their sire by the reversion hypo- thesis, and I have made a number of experiments with the object of testing the probability of this explanation. Some of these experiments I shall now mention.
(1) Pigeons. - With the numerous varieties of pigeons a countless number of experiments have been made by Darwin, Tegetmeier, and other naturalists, and by a host of fanciers. These experiments are especially interesting to biologists, because they point to the conclusion that the crossing of extreme forms leads to reversion towards a remote ancestor - in this case to the wild rock-pigeon, Columba livia. But in addition to this a number of other deductions have been made. It has been inferred, e.g., that it is extremely difficult to eradicate blue when it appears in any strain, and that some breeds are more pre-potent than others.
As I shall have to refer frequently to the wild rock-pigeon, I may here quote Darwin's short but very accurate description of its colouration. He says, "The wild rock-pigeon is of a slaty-blue colour; the wings are crossed by two bars; the croup varies in colour, being generally white in the pigeon of Europe, and blue in that of India; the tail has a black bar close to the end, and the outer webs of the outer tail feathers are edged with white except near the tips." [ 'Animals and Plants,' vol. i, p. 204.]
Darwin found that when different varieties were crossed, the offspring often in the colour of the wings and tail or in some other respects resembled the blue rock, and in one instance by crossing a barb-fantail female with a barb-spot male he produced a bird "which was hardly distinguishable from the wild Shetland species." It only appeared to differ in having the head tinted with red, and like the stomach of a paler blue colour. ['Animals and Plants,' vol i, p. 210. ] Weismann, in referring to this experiment, says, "These breeds [the barb, fantail, and spot], as is well known, differ from the wild pigeon in colour as in many other details, such as the length of the beak and number of tail feathers; and it would therefore be interesting to ascertain whether these racial characters had all disappeared in the grandchild, and had been transformed into the corresponding characters of the wild species. Were this so the reversion might be considered complete. . . . Unfortunately Darwin leaves this point untouched, as he devoted his attention chiefly to the colouration of the species. It seems to me to be very probable, however, from several of his statements, that this was also essentially a mere case of reversion as regards the colouration of the plumage. . . . The other racial characteristics do not at any rate exclude the possibility of a blue colouration; and thus, on the other hand, reversion to the blue colour is not necessarily accompanied by a reversion to all the other characters of the ancestral form." ['Germ-plasm,' pp. 323, 324] I think, though Darwin does not definitely make the assertion, almost complete reversion in his barb-fantail-spot cross is implied. Had this cross in its beak or legs, or length of wings, differed decidedly from the wild rock standard, the fact would most probably have been men-tioned.
Whatever uncertainty remains as to the extent of reversion in pigeons should, however, be dispelled. It is partly with this object in view, and partly to throw new light on inbreeding and prepotency, that I have recently been experimenting with pigeons. Darwin, when dis- cussing prepotency in the pigeon group, says, "In making reciprocal crosses between pouters and fantail pigeons [i.e. crossing them both ways] the pouter race seemed to be prepotent through both sexes over the fantail. But this is probably due to weak power in the fantail, rather than unusually strong power in the pouter; for I have observed that barbs also preponderate over fantails. 'Animals and Plants,' vol. ii, p. 41. But not only are some varieties said to be more prepotent than others, it is generally assumed that certain colours and markings are latent, and ever ready when there is any disturbance of the balance to reassert themselves. To use Mr. Wicking's words, "when a blue or a black and chequered bird, having black wing bars, once appears in any race and is allowed to breed, these characters are so strongly transmitted that it is extremely difficult to eradicate them." ['Animals and Plants,' ' vol. i, p. 210.]
It thus appears that, in addition to settling whether there is complete or almost complete reversion in form as well as in colour, there is the question as to whether the blue colour and the characteristic bars can be easily eradicated; and the further question, can the prepotency of any given breed be increased? can a fantail, e.g., be made prepotent over a pouter?
[* I need not say that in experiments of this kind the desired results, if they come at all, seldom come at once. Even in the most expert hands there is a large percentage of failure, but happily from failures something can generally be learned. ]
Let me deal first with the colouration question. Can the blue colour be altered, and the dark wing bars be eradicated? Fanciers, as stated above, seem to agree that while in certain breeds wing bars may not appear for many generations if pure-bred birds are used, they almost invariably come to the surface when intercrossing is resorted to; and they also believe that when once established it is a difficult matter getting rid of them.
As it happens, I soon succeeded in getting birds without wing bars by making crosses, which could hardly a priori have been expected to give positive results. I crossed a well-bred dark blue fantail, having all the characteristic bars of a rock-pigeon, with a less well bred fantail, also blue, with the exception of the croup, head, and tail, in which there were a number of white feathers.
On two separate occasions these blue fantails produced a well-formed absolutely white fantail. I believe this is an instance of partial reversion, the explanation being that the white offspring took after a white parent of their sire. Fantails are an old breed, and white fantails are especially common. However unlikely the result may to some appear, I was quite prepared for it from what I had already learned about reversion and inbreeding. I next crossed a white fantail cock (which I believed to be inbred) with a blue pouter hen. According to the prevalent view the offspring should have taken after the blue pouter at least in colour and markings, but as it happens the cross-bred bird - there was but one - is almost as white as the fantail, while in form it closely resembles the blue pouter. I see no evidence of reversion in this instance, nor yet of prepotency of the male over the female. It is a case of each parent handing on its most fixed individual characters, the fantail giving the colour, the pouter the form and disposition. The white fantail cock, which in colour proved prepotent over the blue pouter, was next mated with a cross between an owl and an archangel. The archangel hen was a very good example of the copper-coloured variety, the sides of the wings and tail being of a bronzed black hue, and the peak or crest well developed. The owl belonged to the powdered blue English variety. It is small and compact, with a very short "owl-like" beak, rounded head, prominent frill, and short legs.
The owl-archangel cross is far more an owl than an archangel. There is, in fact, nothing either in the colour or form to suggest the archangel parent, and though evidently related to an owl it differs in having the head and beak elongated, in the length of the legs, the absence of a frill, and also in having the wing bars less distinct, and the wing coverts chequered with brown. In all its movements it is nearer the blue rock than an owl, and, as in the blue rock, there is a white croup and twelve tail feathers, the outer one at each side edged with white except near the tip. The head, tail, and croup approach the European blue rock in colour, but the breast and wings are tinged with brown.
The fantail with which the owl-archangel hen was mated is absolutely white, has thirty feathers in its tail, and is sufficiently prepotent to produce an almost white bird with a blue pouter. When the feathers eventually appeared on the offspring of the white fantail and the owl-archangel I was not a little surprised to find that both young birds were blue, one of them almost identical with a young wild blue rock-pigeon.
The wild blue rock varies considerably in colour. In Shetland, where little, if any, intercrossing with the dovecote pigeon has taken place, the croup is as a rule white, and the wings in front of the bars of a uniform light blue colour. In India the croup is generally blue, while the wing coverts are chequered with black. In Madeira, through abundant chequering the front part of the wings often looks almost black. In one of my young birds the croup and wings agree with the Shetland rock- pigeon; in the other the croup, except near the root of the tail, is blue, and the wing coverts are chequered. They thus may be said to illustrate the whole range of variation in the colour of the croup and wings in the wild rock-pigeon of Europe and Asia. The bird with the white croup has several white feathers about the head, and it thus departs from a typical wild bird; but the one with a blue croup is in its colouration the image of a chequered rock-pigeon.
In the bird with chequered wings we have complete reversion, not to the blue rock of Europe so much as to the blue rock of India. The explanation of this may be that the ancestors of both the fantail and the owl were Indian, or at least Eastern birds. If this is so the reversion in the case of the darkest bird is all the more complete and remarkable.
Having referred to the reversion in colour, the question remains, is there reversion also in form? Weismann, it will be remembered, took for granted that Darwin's fantail-barb-spot cross was "a mere case of reversion as regards the colouration of the plumage," and pointed out that "reversion to the blue colour is not necessarily accompanied by a reversion to all the other characters of the ancestral form." In the case of my most typical bird there is, as far as an external examination can show, practically complete reversion. In its measurements it is relatively almost identical with a typical Shetland blue rock. I say relatively because, having only reached maturity, it is not yet full-grown. In its extreme length it is 2 per cent, shorter than a full-grown Shetland bird, and this amount of difference may be said to obtain for the legs, beak, toes, &c. In its attitude and movements it resembles the wild rock-pigeon. If there is auy difference it is in the tail. There are the typical number (twelve) of tail feathers, and the outer margin of the outermost feather at each side is edged with white, except near the tip, but the feathers at each side of the middle line have their inner edges very slightly tilted upwards. This is the only hint of the highly specialised nearly upright fan-like tail of the white sire.
From these experiments with pigeons it is evident that the reversion may be partial or all but complete in form as well as in colour. In the case of the white fantails there is partial reversion in colour only; in the case of the white fantail and the blue pouter - two well-marked and long-established breeds - the struggle amongst the parts ended in the production of a bird taking after the one parent in its colour and the other in its form; with the owl and archangel there was considerable reversion towards the blue rock, but, perhaps because the archangel is a comparatively recent production, none of its characters could be detected in its cross-bred offspring; in the case of the white fantail and owl-archangel cross there is practically complete reversion in form as well as in colour.
Perhaps I may here say that reversion is more a negative than a positive influence, that if I understand it aright complete reversion is mainly due to the development being abruptly arrested so as to reproduce a lost ancestor. Sometimes several printings are required to produce a coloured plate. Were one or more of the printings omitted a kind of "reversion" would be the result. When the owl and archangel are crossed the latest colours added by the fancier are not reproduced, and the older and simpler colours are again made visible. When the white fantail and owl-archangel were crossed all the recently acquired colours were lost, and the common ancestor of all our domestic pigeons was once more re- produced. How the ancestral units of germ-plasm overcome during development the less stable units representing more recently acquired peculiarities can only be guessed at.
It will be remembered that Weismann thought it very probable that Darwin's barb-fantail-spot cross was "essentially a mere case of reversion as regards the colouration of the plumage." In my owl-archangel-fantail cross there is not only more complete reversion as regards the colouration than in Darwin's cross-bred bird; there is, as far as external measurements can show, practically complete reversion as regards form, but more remarkable still, there appears to be reversion as regards the constitution and habits.
A successful and experienced breeder of sporting dogs recently told me that close inbreeding for three or four generations leads to marked senile degeneration. If, however, there are any puppies in a grossly inbred litter that take after a good ancestor several generations removed, they invariably prove the strongest and best. In the same way it is said that if an inbred sire and dam produce a striped dun-coloured foal, it almost always turns out well, and alike in hardiness, staying power, and durability eventually surpasses closely related offspring of a grey, chestnut, or bay colour. That highly bred, not very fertile, delicately nurtured parents, with weak constitutions, should sometimes give rise to vigorous prolific hardy offspring may seem impossible, and yet a great many facts might be marshalled in sup- port of this assumption. Reversion, in fact, seems to lead to a form of rejuvenescence - due presumably to the ancestral units overcoming and controlling the more recently evolved and less stable units, which, if allowed to have their way, would give rise to offspring bearing all the marks of decadence that characterised the immediate ancestors.
My "restored" blue rock looks as vigorous and hardy and compact as a wild bird, and he is greatly admired by fanciers alike for his carriage, form, and colour. He seems to prefer sitting, when off duty, in a dark recess to sunning himself, and alike in his movements and disposition he differs from his companions. He is shy, active in his movements, and when disturbed seems to be not a little concerned about the nest in which his hopes are at present centred.
2. Fowls. - It is generally believed that the game breed of fowls has sprung from one or more varieties of the jungle fowl (Gallus bankiva), and that from the game all the other domestic fowls have been derived. If this is the case, and if there is any truth in the reversion dogma, fowls when crossed should sometimes take on the characters of either the game or the wild breed.
An unusually dark red-breasted game bantam hen was crossed with an Indian-game-Dorking. The result was nine chickens, six o£ which resembled Dorkings, while three in their form and colour resembled game birds. Two of the three, now they are full-grown, only essentially differ from certain varieties of the wild fowl in having a small double instead of a small single comb - the double comb having been inherited from the cross-bred game-Dorking cock. These game-like, cross-bred birds are especially interesting because they are very shy, and fly about like wild birds. There has thus, in this case, been a marked reversion, not only in form and colour, but also in disposition and habits - a not uncommon thing in cross-bred animals.
3. Dogs, - All the breeds of dogs are probably genetically related to the wolf, yet I never heard of a cross being obtained that as closely resembled the wolf as cross-bred pigeons occasionally resemble the wild rock-pigeon. This is perhaps partly because the wolf is not the only wild ancestor, and partly owing to the majority of the most familiar breeds being inbred. All that can be expected of dogs is that they should revert to one of the less remote ancestors, just as the offspring of the blue fantails reverted to their comparatively recent white ancestor. When colour alone is considered, the Dalmatian breed may, I think, be looked upon as the most highly specialised; to wolf-like dogs it stands in the same relation as the zebra does to dun-coloured horses. Dalmatians have probably sprung from pointer-like progenitors, and being comparatively uncommon they are as a rule inbred. Hence, from an extreme cross with a Dalmatian, instead of wolf-coloured or even whole- coloured offspring, we should expect the progeny to approach in colour the Dalmatian's less remote ancestors, i.e. we can only look for partial reversion. I crossed a Dalmatian (one of the lighter varieties) with a well-bred sable collie, and obtained three pups. The pups (which died when quite young and were preserved in spirit) neither resemble young collies nor young Dalmatians, but rather suggest young pointers or foxhounds. The ground colour is nearly white in all three; in two there are five large dark brown blotches, in one four lemon-coloured patches. This seems to be a case of reversion towards the mediaeval ancestors of the Dalmatian.
4. Rabbits. - During the present summer I have bred a large number of white rabbits by mating ordinary white does with a smooth-coated buck. With two exceptions all the young have resembled their parents. The one resembles an Angora, the other as closely resembles a Himalaya rabbit. The Angora might be looked upon as a "freak;" or seeing that it is the only one out of well- nigh fifty rabbits by the white buck, it might be accounted for by saying the doe must have numbered a long-haired rabbit amongst her ancestors. The real explanation doubtless is that the sire of this exceptional rabbit was bred from an Angora rabbit, i.e. the long-haired rabbit is, as far as external characters go, an exact reproduction of its paternal grandmother. The restored "Angora" is not so much interesting because the reversion is, as far as it goes, complete, but because only one out of nearly fifty rabbits by the same buck showed any signs of reversion. I know of a similar case of reversion in the human family. A man, decidedly lighter than an octoroon, married a fair English woman. Of their two children, one is like the mother, the other is quite as dark as a mulatto.
The "restored" Himalaya is even more interesting than the Angora. It exactly reproduces the maternal great-grandmother. But as one of the sisters of the sire had a dark muzzle it is conceivable some of the Himalaya blood came from the paternal great-grandfather. Many other instances of reversion in rabbits might be given, some of which exactly agree with cases of reversion commonly supposed to occur in the human family. These rabbit experiments lend powerful support to the view that a child may be the image of its grandmother, or of its great-grandmother, while the new foal of the Iceland skew-bald pony, to be further referred to, affords very strong testimony in favour of the belief that a child may be the image of its own mother.
That only one of the fifty descendants of the white buck took after his dam (i.e. after their grandmother), while all the others but one resembled his sire (i.e. their grand- father), shows that experiments of this kind must be conducted on a fairly large scale to be of any use. The maternal grandparent was probably represented by germinal units in all the fifty descendants of the white buck, but only in one were they sufficiently potent or sufficiently plentiful in the ripe male germ-cell, or sufficiently lucky to obtain complete control during the co-mingling of the units of protoplasm that accompanies and forms the essential part of fertilisation.
Two reflections that flow from a contemplation of the solitary "restored" Angora may, though somewhat out of place, here be mentioned. The first is, had the doe of the solitary young Angora been previously mated with an Angora buck, she would probably have been cited as a case of "infection," provided of course the breeding of the buck had not been recorded. The other reflection may take the form of a question. If only one out of fifty (i.e. 2 %) of the offspring take after the grandmother, how many of the offspring (supposing there is such a thing as telegony) might be expected to take after the previous sire, whose influence must presumably count for infinitely less than that of an ancestor only one generation removed? I think the answer might be, at least for rabbits, not more than 1 per cent, of the offspring. It will be remembered that Romanes in describing telegony says, "It is so rare that I doubt whether it takes place in more than 1 or 2 per cent, of cases;" adding, however, that "nearly all my professional correspondents would deem this an absurdly low estimate." Evidently, should the fact of telegony be proved, it will be necessary to determine as accurately as possible whether it is a very rare or a comparatively common phenomenon.
5. Ducks. - Another interesting case of reversion may be mentioned. A cross between a common wild duck and a black Cayuga drake produced during the present summer to a common wild drake seventeen ducklings. Of these nine are like the black Cayuga grandsire, while eight take after the wild drake - the immediate sire.
[* To Alexander Cowan, Esq., of Woodslee, Penyeuik, I am indebted for the information about the ducks, and for various other friendly deeds.]

6. Horses. - I may here mention several instances of apparent reversion in the Equidae.
(a) Teeth. - The ancestors of the horse had seven cheek teeth - four premolars and three molars - on each side of the upper jaw. The recent horse has, as a rule, only three (2 - 4) premolars. When a fourth (the wolf-tooth) is present it is small, and usually soon disappears. In zebras there are often four premolars in the upper jaw, and the first is sometimes nearly as large as in the extinct three-toed horse Hipparion. Two years ago I bred a bay Shetland foal, having several distinct stripes on the neck and shoulders, bars in the vicinity of the "knees" and hocks, and, in addition, a distinct dorsal band. In this foal the first premolar (wolf-tooth) was large - relatively nearly as large as in Hipparion.
(b) Digits. - The biologist is as satisfied that the horse has descended from five-toed ancestors as the astronomer is that the earth moves round the sun, and he knows that not very long ago, geologically speaking, all horses had three complete digits encased in three separate hoofs. The digits borne by the fore-limb corresponded to the fore, middle, and ring fingers in man, the hind digits to our three middle toes. As I pointed out some years ago, the horse is still for a time tridactylous. ln a four- weeks horse embryo there are no rudiments of digits in the paddle-shaped limbs, but in a five-weeks embryo there are rudiments of three digits, and at six weeks the foot is a miniature of that of the rhinoceros. [* the horse, hence, does not during development, as some imagined, pass through a five-toed stage, i.e. it is never, even when a very small embryo, pentadactylous like its supposed remote ancestor Phenacodus, in which the skeleton of the fore-foot is, in many ways, wonderfully like that of the human hand.] The second and fourth digits in the embryo horse, though small, are almost as complete as in Hipparion, the three- toed fossil horse preserved in large numbers in the Pikermi beds near Athens. As in Hipparion, the second and fourth digits are asymmetrical. They thus, while forming a nearly symmetrical pair, differ from the large symmetrical middle digit - the only complete digit in recent Equidae. Occasionally a foal is born with two hoofs on one or more of its limbs; at very long intervals a foal appears with three hoofs on one or more of its limbs. Alexander's Bucephalus, e.g., was polydactylous, as was Caesar's favourite horse. I have in my possession four specimens showing extra digits in the horse. Polydactylism is not uncommon in man, and it seems to be still more common in the pig. In man the extra digits seem to be always due to the splitting either of the thumb or of one or more of the fingers. In one of my polydactylous horse limbs the extra digit is without doubt due to the splitting of the third or middle digit. The extra digit in the three remaining specimens is, however, not so easily accounted for. When the large middle digit splits, the two resulting digits are almost identical in form if not in size, in my most complete specimen one of the digits - the inner - is not only very much smaller than the other; it, like the inner digit in Hipparion, is asymmetrical, and the articulation between the first and second phalanges (the first and second pastern-bones) has been obliterated. The smaller inner digit is thus far from being an image of the large functional one; hence it does not seem to have been formed by splitting or dichotomy. But if the extra digit has not in this case been split off from the large middle digit, its existence can only be accounted for in one of two ways. It is either a sport - an instance of abrupt or discontinuous variation - or a restoration of one of the digits found in the three-toed ancestors. Seeing that the horse embryo starts with three toes, and that the lateral digits persist in a nearly perfect though minute form for several months, it seems to me more natural to account for the occasional presence of an asymmetrical extra digit, such as occurs in some of my specimens, by the reversion theory than by any other. As it happens, in the inner (second) digit of the embryo horse I especially examined some years ago the first and second phalanges had all but united. Had this embryonic digit been favoured from the first by an unusual amount of nourishment, it might have increased to form a large extra digit instead of degenerating to form a mere vestige (the "button") at the end of the inner "splint" bone. If in some cases the extra digits in the horse correspond to digits which persisted in the horse family well through the tertiary period, these digits form a very striking instance of reversion.
(c) Forearm. - Keeping to the fore-limb, still another case of reversion may be mentioned. In the ancestors of the horse, as in ourselves, there were two distinct bones in the forearm, the radius and ulna. In all the works dealing with the skeleton of the horse the ulna is described as incomplete. It is said to terminate in a slender process some distance from the lower end of the radius. In very young horse embryos I find the tibia is not only as complete, but nearly as large as the radius. But it is often complete, though not entirely ossified, in foals, and occasionally complete in the adult. In the skeleton of the horse placed by Sir William Flower in the entrance hall of the Natural History Museum, London, there is a complete ulna, and in all horses the lower end of the ulna persists and enters freely into the wrist (" knee") joint. In having at times a complete ulna in the horse we have another instance of reversion.
As the whole question of reversion involves a general acquaintance with animal pedigrees, I may, before referring further to my horse embryos, say a few words about what is usually known as the Recapitulation Theory. In the making of a breech-loading hammerless gun, a piece of wood, is gradually fashioned into a stock, and various kinds of metal are manipulated in diverse ways to form the lock and barrels. From the first it is evident what the workmen are about. But if a gun were constructed on the plan followed by nature in the making of a mammal, the procedure would be very different. The gun-maker (who would require to be a miracle worker as well) would go to work in some such roundabout way as the following. Starting with a piece of clay, he would set about transforming it into flint, but before completing the process would convert it into something remotely resembling a boomerang or a knob-kerry. By further legerdemain the "throwing stick" would become a cross-bow, and this, by a long and intricate series of transformations, would assume, or all but assume, in turn the characters of a flint gun, various kinds of muzzle- loaders, and breech-loaders with hammers. Eventually it would take the form of a hammerless ejector.
In no Vertebrate is the development direct, however rapid it may be. For example, the horse, an all but invisible speck of protoplasm to start with, passes at the outset through a series of remarkable changes (which so far can only be guessed at), to emerge at the end of the third week as a somewhat fish-like creature almost bent double over a large active heart. The tail is bilobed, like that of a mermaid (manatee), but neither limbs nor jaws have yet appeared. A mammal at no stage can make use of gills, yet the horse embryo has, for a time, three pairs of gill pouches. A quarter of an inch long at three weeks, the horse embryo is half as long again at four weeks. It might, at this stage, pass for a human embryo of a similar age. There are paddle-like limbs, and a lizard-like tail many times longer than the limbs. At five weeks the limbs are still paddle-like, but in each paddle the foundations of three digits (2, 3, and 4) have appeared. A week later the fore-limbs are longer, have undergone rotation inwards, and the three digits in each are more distinct. The jaws have now appeared, and there is no longer any indication of gill pouches. In the six-weeks horse (though under three quarters of an inch in length) we have a partial restoration of the earlier three-toed ancestors of the Equidae, but ere the seventh week is reached all external indication of the outer (2 and 4) digits has vanished, and the single remaining digit (3) is beginning to assume the characteristic equine form. By the end of the eighth week, though the embryo is barely two and a half inches in length, we have in many ways a miniature horse, the limbs extended as if ready for action, and the tail only reaching the hocks. If at six weeks the cross-bow stage has, as it were, been reached, by the eighth we have got as far as a flint gun.
As it happens, these and other facts to be mentioned presently have a bearing not only on the reversion theory, but also on a subject attracting much attention at the present time, viz. the origin of the three great groups of Mammals. During recent years this subject has been often discussed, and we seem to be drifting further and further from the view that the higher Mammals (Eutheria) have descended from the Marsupials - the opossum and kangaroo group, - while they in their turn originated from the egg-layng Monotremes, the group now represented by the duck-mole and echidna.
It may be more than a mere coincidence that, for a few days, the horse embryo forcibly reminds one of some of the young Marsupials. In nearly all Marsupials nourishment up to the time of birth can only be derived from food - "uterine milk - filtering into the yolkless yolk-sac. This necessitates premature birth. In the opossum, e.g., the young are transferred to the pouch as soon as they can hang on to the teats. In the higher Mammals and in certain Marsupials a more permanent plan is adopted for the nourishment of the embryo. The embryo-horse, during the first six weeks, is nourished like the young opossum and kangaroo by "uterine milk" filtering into the yolk-sac; but beyond this period the foetal blood is brought into close relation with, that of its dam, and thus a fresh food-supply is tapped. During the seventh week, while the new machinery is being fitted up, the embryo, uncertain as to its fate, behaves like a young opossum when preparing for its life in the pouch. Few now suppose that young opossums, wallabies (rock kangaroos), and other Marsupials reach the pouch by crawling down the teats. On the other hand, few seem to know how the young wallaby, e.g., manages to glue itself so firmly to the teat as soon as it. reaches the pouch. At the end of each teat (as Mr. le Souef of Melbourne recently explained to me) there projects a delicate tube. The female wallaby conveys the young as they are born by means of her lips to the pouch, which is kept open during the process by her short fore-limbs. Even at this early period the lips of the young, when stimulated, may contract; at any rate, the minute tube hanging from the end of the teat gets into the mouth of the helpless youngster, and is probably held in position for a time by the lips. The lips, however, if active at this stage, would soon get exhausted. To prevent the young one slipping, the slender tube at the end of the teat is distended, it is dilated so as to fill the mouth, the result being that the young are so firmly fixed that a considerable force is required to drag them from their moorings, and when once removed it is impossible to replace them. The dead or helpless embryos, sometimes found lying in the pouch, have probably either slipped from the lips of the mother or dropped from the teat before the delicate terminal tube was inflated. In the young Marsupial, it may be mentioned, the lips are relieved from the business of sucking; the milk is pumped into their mouths by certain muscles compressing the mammary glands.
It may seem a far cry from a new-born wallaby to a seven-weeks horse embryo, but the distance may not be so great as it seems. Not only is the lower jaw in a very backward state in the young horse at the end of the sixth week, it is still greatly shorter than the upper jaw at the beginning of the forty-seventh day. Evidently during the last two days of the seventh week the development of the structures around the mouth proceeds apace, as if it were unexpectedly necessary to prepare the embryo for premature birth, and for playing the part of a helpless opossum. By the end of the forty-ninth day the lower jaw decidedly projects beyond the upper, the tongue is well formed and slightly raised from the floor of the mouth, which is sufficiently open to show a fair-sized, well-formed space between its roof and the well-moulded tongue. A point of still greater interest is that both the upper and lower lips are so notched that, if approximated, a gap would be left about the size of the slender terminal portion of the teat in a Marsupial. Another point is that, as the lips rapidly develop, rudiments of the vibrissae (sensitive hairs) make their appearance around the muzzle. As I said before, all these changes taking place about the time the horse would presumably be born were it a Marsupial may be a mere coincidence, and quite meaningless. They may, however, point to a time when the ancestors of the horse were as primitive as some of the Australian Marsupials are to-day, to a time when they were prematurely ushered into the world in a perfectly helpless condition; and they may further indicate that, if the higher Mammals (the Eutheria) have not actually sprung from amongst the Marsupials, they and the ancient Marsupials travelled for a time along nearly parallel lines. If this is the explanation, it follows that at the end of the seventh week (when the horse embryo is little over an inch in length) we have an extremely interesting example of recapitulation, a kind of embryonic reversion. Evidently it is but a passing phase, for ere the eighth week comes to an end the mouth is tightly closed - the lower lip partly overlapping the upper at each side - and the eyes are also closed. The eyes, I might have mentioned, are widely open during the whole of the seventh week.
Before leaving the subject of reversion I may refer to a hinny (jennet) recently added to my stud. This young hinny (a cross between a bay Welsh pony stallion and a common ass) is of a yellowish-brown colour - the colour of the wild horse of the Dzungaria Desert - and has distinct bars at the "knees" and hocks, as well as a dorsal band and a shoulder stripe. The leg bars are evidently due to reversion, as no leg bars are present in either of the parents. Reversion of this kind is very common in mules, and is not unknown amongst pure-bred asses and zebras. A zebra foal, e.g., born during the present summer in the Regent's Park gardens, London, is striped to the hoofs, and is in other respects like the zebra named after Mr. Selous. In the parents, which belong to the Chapman variety, the legs are only partially striped. Several of my hybrids have more bars on their legs than their sire.
From what has been said as to reversion, it is evident that breeders should in all cases direct as much if not more attention to the ancestors than to the parents of their breeding stock, that they should expect and make allowance for reversion, for "regression towards mediocrity," especially remembering that the more unlike the parents the greater is likely to be the reversion in the offspring, and that the latest acquired peculiarities are likely to go first unless they have been "fixed" by inbreeding, or are of the nature of "sports." [* Evidently the more inbred and closely related the parents the more limited will be the range of reversion, but still even in grossly inbred stock reversion will occasionally occur, especially to prepotent and not very far removed ancestors.]
As indicated in the last of the three papers, it has been the fashion for some time to throw doubt on the reversion hypothesis. It has been stated that " around the term reversion a singular set of false ideas have gathered themselves." [Bateson, 'Materials for the Study of Variation,' p. 76] But statements of this kind do not disprove reversion. Even if the hypothesis in question has been invoked to account for the appearance of new forms which were either sports or the product of discontinuous variation, it does not follow reversion never occurs. I think it must be admitted the pigeon and rabbit experiments go a long way towards proving the fact of reversion, and the more I contemplate my zebra hybrids the more convinced I am that they are neither new creations in the strict sense of the terra, nor yet intermediate forms; and if they are neither the one nor the other, they must be more or less accurate restorations of their comparatively remote ancestors. The heredity problem is sufficiently difficult as it is, but if we are debarred from invoking the assistance of the reversion hypothesis it will become hopelessly incomprehensible. In conclusion, I may point out that even from a practical point of view reversion is of extreme importance, for it indicates how in some cases varieties and breeds which have through inbreeding undergone senile degeneration may be regenerated without the loss of their best and most prized characteristics.

2. Prepotency and Inbreeding. - More than thirty years ago Darwin recognised that "the subject of prepotency is extremely intricate." It must be admitted this intricacy continues, and is likely to continue until the laws of heredity are better understood. Amongst the other difficulties we have to contend with, there is the impossibility of determining the amount of prepotency in any given race or individual, and the fact that prepotency may altogether escape detection, or exist through gross inbreeding where it is least expected. The leader of any given wild herd may be decidedly prepotent, but unless he is mated with the members of some other herd presenting different characters the prepotency may escape notice; and again, the members of any given section of a species may, through inbreeding, be highly prepotent, while the members of the other sections (owing to abundant facilities for intercrossing) may be non-prepotent. What is true of individuals may be true of races and varieties. The Jews, as a race, are more prepotent than the English - are better or purer bred; but the prepotency only declares itself when intermarriages take place. On the other hand, any given member of a non-prepotent race may, through inbreeding (sib-breeding), be more prepotent than any given member of a decidedly prepotent race. Hitherto prepotency has almost invariably been associated with inbreeding, and many believe not only that it can be induced by inbreeding, but that apart from inbreeding it is impossible. On the other hand, sports and certain marked variations are often prepotent, i.e. they resist the swamping influence of inter- crossing, and hence it would be difficult to prove in many cases that prepotency has not originated with, or is an attribute of, sports. [* A sport is an abnormal or aberrant variety the result of sudden and pronounced variation in one or more directions. The horse compared with a dog, or its old enemy the wolf, is a poor jumper. If a horse were suddenly to appear that could leap nearly twice its own height it would be a sport, as would be a horse with extremely short legs or without a tail. An upright mane might either be a sport or the result of reversion, while a hairless horse would be considered a sport. ]
Mr. Galton has recently gone further than this, and given it as his opinion that prepotency is, in itself, a sport; to use his own words, "high prepotency does not arise through normal variation, but must rank as a heritable sport or aberrant variation." [Nature, July 14th, 1898.] That sports are frequently prepotent has been placed beyond doubt, but that prepotency is in itself a sport has not yet been proved. When we are able to account for prepotency, why any given variety is able to withstand the swamping influence of intercrossing, we shall, I think, be nearer an explanation of the origin of species.
Mr. Galton, in his suggestive paper on prepotency, says nothing about interbreeding. Perhaps, while believing "high prepotency" rapidly acquired is due to an aberrant variation, he will be prepared to admit that a lower grade of prepotency may be induced by inbreeding. I believe inbreeding is common amongst wild animals, and that by inducing prepotency it plays an important part in the origin of species. Variation is of fundamental importance; the destruction of the unfit is also important; but simple variation and the survival of the fittest, unless sterility or prepotency is implied, are not sufficient to account for the origin, from a few simple forms, of a countless number of plants and animals, many of them amazingly complex in organisation. The need of something above and beyond variation and natural selection was especially recognised by the late Mr. Romanes. In what he termed physiological selection he believed he had found the additional factor, physiological selection being something by which isolation was secured, which made sure that some of the varieties so abundantly produced in nature would have a chance of perpetuating themselves, i.e. escape being swamped by intercrossing. In the case of the domestic animals the breeder plays the part of the additional factor; by isolation he prevents intercrossing. Without fences the famous breeds of cattle that have been so carefully built up would, unless highly prepotent, soon cease to exist as distinct breeds, and would ultimately be merged through intercrossing into as many varieties as there are distinct or isolated areas.
Hitherto the explanation of one variety persisting while others, equally fit in every way, vanished, has sometimes been that the members of the more fortunate variety, while -fertile with each other, were sterile, or at least less fertile, with other varieties and with the parent species. It is, of course, conceivable that some sports are sterile except amongst themselves, just as some sports are prepotent, but satisfactory evidence of this is still wanting. The explanation which prepotency affords does away with the need of sterility, and it does away with the need of rigid isolation, with natural barriers or fences, because the prepotent forms have so much "character" that, however mated, some of the offspring inherit their own structural and other peculiarities.
What, it may here be asked, do we understand by prepotency and interbreeding? Any animal, male or female, which strongly impresses its own peculiarities of form, colour, disposition, &c., on its offspring, is prepotent, while animals that are the offspring of more or less intimately related parents are interbred, - when the parents have been closely related for several generations they g.re said to be grossly inbred. There is, however, nothing absolute about either prepotency or inbreeding, they are relative terms. An animal (male or female) may be prepotent in some respects and not in others (and this whether the prepotency has been acquired by inbreeding or through a "sport"), or prepotent with one mate and not with another, prepotent one year but not the next, because prepotency is of necessity subject to the influence of variation and reversion, and also doubtless to nutrition - more especially of the germ-plasm prior to fertilisation. Prepotency, when associated with useful characters, is highly beneficial, while inbreeding is often a doubtful heritage. Nature only tolerates inbreeding up to a certain point, for while it may assist in perpetuating useful characters by inducing prepotency, it often does this at the expense of vitality - it may be of fertility as well. It is conceivable that inbreeding has played an important part in the extinction of species; it has undoubtedly been the means of deteriorating, if not actually destroying, many of the breeds and varieties artificially produced. A carefully conducted series of experiments, showing the effects of inbreeding on various kinds of animals, is still greatly needed. When such a series is forthcoming it will be time enough to discuss at length the physiology of inbreeding. Still, something further may well, at this stage, be said about inbreeding and prepotency. It will, I think, be admitted that prepotency may be due to either natural or artificial causes, or be partly due to both. In nature, prepotency may (1) arise spontaneously and abruptly along with sports in one or more directions, or gradually with the help of natural selection - whether it may arise as a special variation apart from a sport, as Mr. Galton seems to think, I am not prepared to say; (2) it may be gradually acquired when a few individuals of any given species or variety are so isolated that inbreeding is inevitable. Artificial prepotency may be slowly induced by inbreeding, but I doubt if prepotency can be rapidly produced artificially apart from inbreeding. It may therefore be taken for granted that interbreeding (1) induces prepotency by fixing the characters of the particular variety selected; (2) in as far as it prevents or at least limits intercrossing it diminishes the chance of reversion on the one hand, and restricts the range of variation on the other, thus tending to maintain undiluted the distinctive characters of the type. But it must be added, inbreeding, while keeping the "blood" pure, tends to weaken it by diminishing the vitality of the breed. Prepotency, on the other hand, however acquired, alike in nature, on the farm, or in the dovecote, tends to arrest or neutralise the swamping effects of intercrossing. Some breeders say they can produce a horse so prepotent, so fixed by interbreeding, that it will produce its like however mated. A famous breeder of high-class ponies once boasted that he had a filly so prepotent through inbreeding that though she was sent to the best Clydesdale stallion in Scotland she would throw a colt showing no cart blood - provided the Clydesdale was not also the product of inbreeding. Nature might make a similar boast, the essential difference being that while in the one case misfits and "weeds" are usually preserved, in the other they are ruthlessly destroyed and buried out of sight - in the struggle for existence the inbred "weeds go to the wall.
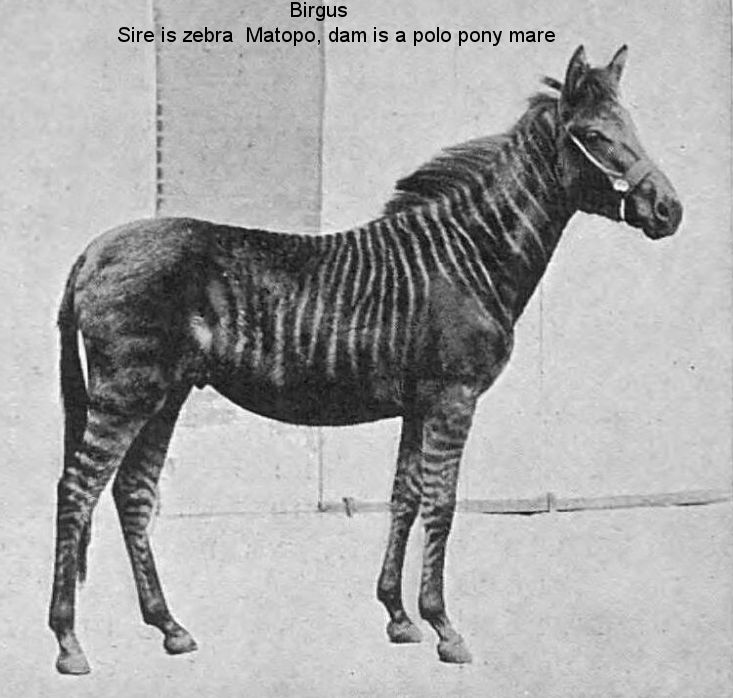
The careful observer soon recognises that all through animated nature the greatest effort is made to secure rejuvenescence. In the animal kingdom, and largely in the vegetable also, this renewal of youth, without which vigorous life is impossible, is secured by intercrossing or cross-fertilisation." [* There is obviously no real difference between cross-fertilisation and intercrossing. Whether we interbreed or intercross, engage in " line " breeding or "cross" breeding, we are making use of cross-fertilisation. Further, I may add the difference between intercrossing and hybridising is one of degree, not of kind.] The ingenuity displayed by plants and animals to secure cross-fertilisation is simply marvellous. What a different world ours would be Lad it been peopled from first to last with sexless forms, with millions of virgin females! The antithesis of intercrossing (cross-fertilisation) is interbreeding (inbreeding). Hence when we grossly inbreed we do our utmost to arrest nature's supremest effort; for though all the elaborate and intricate mental and physical processes are complied with when close inbreeding is practised, there is little or no real intercrossing. In inbred herds, e.g., the males and females are so like each other that the offspring, were it possible, might almost as well be raised by buds or cuttings. In inbreeding the letter of the law is observed, but not the spirit. In proof of this take the case of the celebrated one thousand guinea bull "Comet." The 'Herd-book' tells us that "the bull 'Bolingbroke' and the cow 'Phenix,' which were more closely related to each other than half-brother and sister, were coupled, and produced the bull 'Favourite.' 'Favourite' was then coupled with his dam, and produced the cow 'Young Phoenix.' He was then coupled with his own daughter ('Young Phoenix'), and their produce was the world- famed 'Comet.' " It must be admitted there was very little intercrossing in the case of "Comet," there was still less in "Clarissa," the dam of the celebrated cow "Restless," and, as a matter of fact, many breeders do their utmost to do away with intercrossing; they act as if they thought nature in introducing intercrossing had blundered. This applies to fanciers and to breeders of horses and sheep as well as to breeders of cattle. A writer in the Field of April 9th, 1898, tells us he had heard "Mr. Joseph Osborne, the ablest authority living on British thoroughbreds, declare that you cannot now get too much of Birdcatcher." But cross-fertilisation, it may be mentioned, not only serves to maintain the organs and tissues of plants and animals at a high state of efficacy; it fosters if it is not the prime cause of variation. It is conceivable in some planets less favoured than our own the plan of cross-fertilisation has not yet been arrived at, or, in transcendental language, has not yet been introduced. Chaos may still reign in certain parts of the universe, and instead of a great variety of plants and animals there may be but a bathybial-like layer of protoplasm - a rind of living seething slime, or, at the most, countless numbers of simple unicellular organisms. In such a case the introduction of cross- fertilisation or its equivalent would work miracles more wonderful than the making of dry bones live. In the case of our own planet untold ages may have elapsed before the raw material of life was capable of benefiting by the elevating influence and resuscitating power of rejuvenescence or cross-fertilisation.
But while exalting the influence of intercrossing - one of the most potent and far-reaching of all the vital forces in nature - it is well to remember that, as already hinted, by intercrossing alone, even with the aid of natural selection- with some means of getting rid of the " unfit" or useless variations - the conditions which now prevail in the animal and vegetable kingdoms could never have been reached. Interbreeding (inbreeding) may have ably seconded intercrossing in the origin of species. For though inbreeding in excess saps the life-springs, and leads to decline, degeneration, and death of the species, perhaps, as well as of the race, it may (under the control of natural selection) be the means of giving certain types or varieties a chance of surviving, - in a word, inbreeding may counteract the levelling, if not regressive tendency of cross-fertilisation - may arrest as with a wall the "swamping effects of intercrossing." As breeders and fanciers select, and with the help of inbreeding fix, from amongst the varieties so plentifully provided, such as they consider most useful or most beautiful, so in nature the varieties that pass in the severe competition that everywhere prevails may be fixed by inbreeding, and have a chance of, in course of time, forming new species. It thus appears that interbreeding may serve as a check to intercrossing, that while intercrossing tends to give a new lease of life, and at the same time favours the appearance of new varieties, interbreeding tends to fix and perpetuate certain varieties, though when carried too far it not only arrests variation, but diminishes vitality and makes for senility and degeneration."
[* Inbreeding is often said to lead to "atavism." If atavism and reversion or regression are considered one and the same thing, then I consider it a mistake to say inbreeding induces atavism. Inbreeding almost invariably leads to degeneration, which is quite a different thing from regression. Cross-breeding, in virtue of reversion, may restore all the vigour and vitality of the lost founders of some of our degraded inbred herds. Neither hybrids nor half-breeds nor mongrels are necessarily degenerate; they are usually intelligent and vigorous enough, while inbred animals may be physically feeble, and mentally on the level of imbeciles or idiots. The individual in an inbred litter of puppies which shows most evidence of reversion is sometimes the only survivor; he is invariably the most vigorous. In the same way, a horse bearing the ancestral colours is often as hardy as a mule.]
It is not hard to understand that a new individual, developed from the union of two germ-cells from differently constituted individuals, should more or less differ from his parents and all their ancestors. Neither is it difficult to understand how the offspring of closely related parents sprung from closely related ancestors should be almost the image of their parents. But it is not so evident why inbreeding should lead to loss of vitality or to senility. The explanation seems to be that the stuff of which animals and plants are made - the living clay or protoplasm - has a limited lease of life, is only capable of giving rise to a limited number of generations unless revived or rejuvenated, unless "fresh blood" is introduced.
Remarkable evidence of this has recently been forthcoming from amongst the Protozoa - the group which includes the simplest and smallest living animals. Some of the infusorians multiply by simple division. Like a single colourless blood-corpuscle, one divides into two, each again divides, and so on the process goes, but not indefinitely. There comes a time when the process of division is arrested, "senile degeneracy" sets in, and the new individuals, incapable of feeding or growing, soon die. If, however, an exchange of protoplasm is occasionally allowed, if what is technically known as rejuvenescence occurs at intervals, infusoria multiply indefinitely; they become, in a sense, immortal. The protoplasm making up the organs and tissues of any two closely related, animals is of necessity almost identical. When such animals are mated cross-fertilisation can hardly be said. to take place, and if this is repeated again and again the individual cells (the germ-cells included) doubtless degenerate in very much the same way as the unicellular infusorians when prevented from conjugating. The result in the highly organised multicellular animal is likely to be the same as in the simple unicellular, viz. senile degeneration of the cells forming or destined to form the organs and tissues.
In these days, when nothing is taken for granted, the baneful influence of gross inbreeding is sometimes doubted, and some are not even satisfied that inbreeding leads to prepotency. It may therefore be well to give some instances of the effects of inbreeding.
Bakewell, Webb, Colling, and Bates were among the first advocates of inbreeding. One of their followers was Amos Cruickshank. After a time Cruickshank discovered inbreeding had been carried too far, his fine herd of cattle was threatened with destruction, and he wisely resorted to intercrossing to rejuvenate his degenerating stock by the introduction of fresh blood. Why a little "new blood," it may be from a herd living under almost identical conditions with the dwindling one, should have so profound an influence has not yet been explained. When we discover why an exchange of molecules between two infusorians at an interval of about two hundred generations is sufficient to arrest degeneration, we shall, perhaps, understand why the occasional introduction of "new blood," or of the same blood from, say, over the seas, may prove the salvation of a herd.
There is abundant evidence that inbreeding leads to prepotency. One or two instances may, however, be given. The Dalmatian dog already mentioned is inbred, as indeed are most Dalmatians. When bred with a pedigree collie, the result was, as already stated, three pups with a white ground colour like the sire, and several large blotches such as presumably characterised the sire's ancestors. In this case the prepotency of the Dalmatian is so marked that the collie can hardly be said to be represented in the offspring. A still better instance of inbreeding in hounds is given by the late Sir Everett Millais. Sir Everett crossed a bloodhound with a typical tricoloured basset dog. The half-breeds were bassets in form, but not quite bassets in colour. When these crosses were bred with a male basset, the majority of the pups presented both the form and colour of highly bred bassets. It may be mentioned that the bassets are bloodhounds with short crooked legs, and that the English bassets have all descended from a few individuals imported from France not many years ago. They are all decidedly inbred. Hence, although the bloodhound used was inbred and of an older type than the basset, the offspring were in form bassets. Evidently Sir Everett believed that had the bloodhound dam been less inbred the crosses would have been from the first bassets in colour as well as in form.
[* Sir Everett Millais, who worked long with highly bred stock, points out some of the evils of inbreeding. He says of inbred dogs, distemper carries off about 60 to 70 per cent, of those attacked, and that in breeding to type we too frequently bring about hereditary deformity, rickets, and other diseases.]
It seems to be generally admitted that British cattle have mainly descended from the ancient Celtic shorthorns, from Chillingham-like cattle introduced by the Romans, and from longhorns imported from Holstein, &c. Whether this be so or not, there is no evidence that any of the less remote ancestors of our modern cattle were hornless; there was a time, doubtless, when the representatives of our cattle, sheep, deer, and antelopes were all hornless, but that was a very long time ago. It follows that the polled cattle of to-day are recent productions; whether they are sports or reversions we need not wait to consider. When long-horned Highland Heifers, such as may be seen any day in large numbers at Poltalloch, Argyleshire, are crossed with hornless Galloway bulls, the offspring are not only hornless, but sometimes so like Galloways that they are with great difficulty distinguished from pure-bred Galloways. This is again the result of inbreeding; the older race counts for nothing in the presence of the inbred black Galloways with no pedigree worth mentioning in the presence of the long-horned Highlanders. When two inbred strains are crossed the result is rather curious; e.g. when white shorthorns and Galloways are crossed the offspring are almost invariably blue greys and hornless: in this case the prepotency of the Galloway, though still predominating, is considerably diminished by the inbreeding of the shorthorn.
The Iceland skewbald pony, already mentioned, is also a striking instance of inbreeding. She first of all produced a light bay foal to a pony of her own kith and kin, - a pony presumably more inbred than herself. Her next foal was the hybrid "Heckla," a dark dun, indistinctly striped, but without the smallest patch of white anywhere. This summer her foal, to a bay Shetland pony," is almost the exact image of herself, not only in make and colour, but in the shape and position of the rather peculiarly arranged yellowish-brown patches. [* To this bay pony three bay mares, had bay foals, and a black mare had a chestnut foal, but a piebald mare had a piebald foal. The Iceland ponies are probably more inbred than the ordinary Shetland ponies, i.e. the ponies not in the Shetland stud-book.] Evidence, apparently trustworthy, of the evils of inbreeding will be found in Mr. Low's great work on the 'Domesticated Animals of Great Britain.' He says a gentleman experimented with foxhounds on a large scale, with the result that through inbreeding "the race actually became monstrous and perished." The same happened with hogs. After a few generations they became diminished in size, the bristles were changed into hairs, the limbs became short and feeble, the fertility was diminished, and the mothers were unable to nourish their young. Low adds, "If the experiment be carried as far as the case will allow, the feeble and frequently monstrous offspring will be incapable of being reared up, and the miserable race will utterly perish." From what takes place with guinea-pigs I think these statements may be at once accepted as trustworthy; nevertheless it is desirable that the experiment with hogs be repeated, and a complete record of the results published.
I am not aware if any corresponding experiments have been made with horses, unless, indeed, the breeding of race-horses may be looked upon us a colossal and costly experiment of this kind. It seems to be generally admitted that English race-horses are inbred. Sir Walter Gilbey speaks of their inherent galloping action and speed as having been implanted "by inbreeding during nearly two hundred years." ['On Breeding Carriage Horses,' 1898, p. 10.] It may, however, be again pointed out that there is plenty of inbreeding in nature, and that the evils we associate with inbreeding may often be counteracted by rigid and persistent selection, by preventing the weaklings from leaving descendants. If in staying power and constitution the modern thoroughbreds are defective, it is not so much that they have descended from a few imported sires, that they "trace their descent to the Byerly Turk imported in 1689, to the Darley Arabian imported in 1710, and the Godolphin Arabian imported twenty years later," as that the physically "unfit" as well as the "fit" have often been allowed to perpetuate themselves, - "blood " and speed only having, as a rule, counted in the selection alike of sires and dams. [Loc. cit., p. 16.] The descendants of the horses that escaped from the Spaniards after the discovery of America were rarely found wanting in hardiness and general fitness, even when small and unshapely; and in the same way the descendants of modern thoroughbred sires now reared under trying conditions in Montana are fit enough, though often not larger than polo ponies. The cold and limited supply of suitable food in winter, or the drought and occasional scarcity of food during summer, as effectively get rid of the weaklings and the overgrown individuals as a prolonged dry season decimates the zebras, antelopes, and other grass-eaters in Central Africa. It is not difficult to realise how unfavourable surroundings would systematically and persistently operate in the prairies and savannahs in almost the opposite direction to that followed by breeders of thoroughbreds. The process of elimination would be mercilessly at work amongst the foals, and proceed without rest or pause during colthood, maturity, and old age.
The foals unable from the first to keep up with their dams would soon perish, the colts that strayed would often have a like fate, while both old and young, incapable of withstanding periods of drought or other unfavourable circumstances, would one by one drop from the ranks, and fall an easy prey to the puma, jaguar, and other flesh-eaters. By the ruthless destruction of immense numbers - not necessarily only the unfit, as we are too apt to take for granted - the old troops would occasionally be broken up, and new and differently constituted troops (including a number of unrelated individuals) would be formed, with the result that the evil effects of inbreeding would be minimised, if not absolutely counteracted. A corresponding destruction of the fit and unfit alike has, of course, been impossible amongst our race-horses during the two centuries that have elapsed since the introduction of Byerly Turk.
I have been recently, in a small way, testing the constitution of various breeds at different ages. Perhaps, the conditions being somewhat unfavourable, the test has been too severe. The plan has been to see how long various breeds may safely be left out night and day in a large field nearly 700 feet above the sea level, but surrounded by woods and sloping on all sides from a central plateau. Large sheds are available, and abundant food is provided. A number of ponies continuously out during last winter (1897-98) did extremely well. Amongst others were Shetland, Iceland, Norwegian, and Highland ponies, a New Forest pony and her two-year-old mule foal, an Exmoor pony, a half-Arab mare, and a small Clydesdale mare. Four thoroughbred mares were rather the worse than the better of being allowed out during the warmer part of the day. A light cream-coloured, half-bred mare, though apparently well in the evening of the 9th of December, was found dead on the morning of the 10th, apparently from a sudden chill. This mare (in foal to the zebra) was in good condition, but, though she carried a fairly heavy coat, she was thin-skinned, - very light duns have probably weak constitutions. During the present autumn (1898) I have had further evidence that the thoroughbred constitution is veiy delicate. Early in August we had three cold, wet days. This was fatal to a zebra-hybrid foal out of a three-year-old, small, thoroughbred bay mare, - none of the other foals suffered from this visitation of the east wind. Later in the season three thoroughbred mares plainly showed the cold was too much for them, and during the last week of October a two-year-old and a yearling thoroughbred completely broke down. So far (November) the three yearling hybrids, the Clydesdale's second hybrid foal, a ten-year-old hybrid out of a zebra mare, a hinny foal (born in July), a yearling out of a New Forest pony by an Arab (Benazrek), an Iceland and a Shetland foal, have all done extremely well, though continuously out night and day. I am inclined to think that under 10 per cent, of thoroughbred foals, and under 20 per cent, of yearlings, would survive if exposed throughout the winter in the vicinity of the Pentlands. In more genial surroundings a larger number might survive, but they would probably never reach a height of fifteen hands.
As a matter of fact, the English race-horse, compared with even the Arab, is like a hothouse plant that only manages to hold its own when forced and nursed with unusual care, and after all, except for covering very short distances at a great speed, the majority of the hundreds annually bred are of comparatively little use. Breeders flatter themselves that thoroughbreds have since 1689 increased on an average eight or nine inches (from 13.2 to nearly 15.3 hands), but they forget this was partly due to the introduction of Arab blood, and that the size of a horse is very much a question of selection, food, and favourable surroundings. If the increase in size and increase of speed have, as is alleged, been accompanied by a diminution in the staying power and general fitness, the gain can hardly be held to compensate for the loss. That there has been a falling off in the thoroughbred may be inferred from "the smallness of the percentage of even tolerably successful horses out of the prodigious number bred at an enormous outlay." [* 'Race-horses,' Sir Walter Gilbey, 1898, p. 6]
Two years ago thirty-two yearlings were sold for 51,250 guineas. "These thirty-two yearlings are represented by two winners of five races, Florio Rubbatino and La Reine, who have contributed about £2000 to the total cost; and there is not, as far as can be known, a single one of the remaining thirty with any prospect of making a race-horse." [Quoted from the Times of December 27th, 1897, by Sir Walter Gilbey, 'Race-horses,' p. 6.]
When it is remembered that in the majority of English races very little staying power is now needed, the failure of the thoroughbred is all the more remarkable. If it is true, as many assert, that the English racehorses are, as a breed, on the down grade, the time for the renewing of their youth has more than come. How, it may be asked, can the needful rejuvenescence be accomplished? Careful selection and careful rearing, on the lines suggested by Sir Walter Gilbey, may do much to arrest further de- generation; but to secure a new lease of life - real rejuvenescence - there are, I believe, only two possible modes of procedure. The one is the introduction of new blood, the other is to induce reversion without introducing new blood. As already mentioned, Millais, when he wished to rejuvenate his bassets, went straight to the original source of the breed, i.e. he crossed his bassets with bloodhounds. If Millais's example - which seems to have produced the desired result - were followed, a number of new unrelated Barb and Arab sires, or rather Barb and Arab mares, would be imported, and put to our best English thoroughbreds. But breeders hesitate in making a fresh start of this kind, for the simple and obvious reason that the English racehorse is not only larger than modern Arabs and Barbs, but also fleeter. Were a number of three or four mile races substituted for some of the shorter ones so much in vogue, the introduction of new blood would become imperative, with the result, doubtless, that ere long there would be rejuvenated horses as large and as fleet as the present thoroughbred; and, in addition, able to gallop as far, and maintain their place as long on the turf, as the giants of bygone days. Failing the introduction of new blood, of say a number of the best mares the desert of Arabia can produce, there is but the alternative of crossing thoroughbreds which have been living under as different conditions as possible, the object being to take advantage of the great good that often results from a complete change of the surroundings or environment. Changes affect all the systems, but most of all the reproductive system: they in some cases arrest degeneration; they may even lead to a kind of rejuvenescence.
In carrying out this limited form of intercrossing, two plans might be followed. In the first place, carefully selected American or Australian thoroughbred mares might be put to equally carefully selected English bred horses having the same form and general characteristics. By this plan the risk of reversion would be reduced to a minimum. In the next place, mares or horses from over the sea might be mated with English thoroughbreds of a distinctly different type, with the view of inducing, or at least courting, reversion. Both plans might give good results, but the second would probably be most effective, more especially if there was reversion to a famous prepotent ancestor, a famous "sport" either in the line of the sire or dam. It would, of course, be necessary to remember that the offspring would tend to take most after the prepotent parent; in many cases the cross-bred stock would, from the first, resemble English thoroughbreds in appearance, though differing from them considerably in constitution. In selecting American or Australian sires or dams, regard should be had to their form and record, and, other things being equal, horses should be selected that are free from blood recently introduced from the mother country, i.e. the purest available descendants of the first good race-horses imported (into the States or Australia) should be preferred.
Perhaps the best way to maintain the staying power and constitution as well as the speed of the English racehorse would be to have recourse now and again to Australia or New Zealand for sires and dams equal, if possible, in fleetness and size to our own thoroughbreds, but differing from them in having a recent dash of Arab blood in their veins.
Having discussed at some length prepotency and inbreeding, various problems may now be considered with which these subjects are more or less intimately involved.
Breeders, to their credit be it said, like biologists, are more or less guided by certain working hypotheses. It is not their blame if some of the tenets making up their creed are found wanting when put to the experimental test, or that some of the explanations of out-of-the-way phenomena are far-fetched or tinged with mediaevalism. The creed of the biologist is ever changing, and, except in the case of mere hodmen, speculation and theory are ever in advance of the facts. But with the breeder, perhaps more than the biologist, superstitions are likely to flourish, for it is beyond the province of the average breeder to test his beliefs by systematic experiments. There is not yet a Newmarket confession to which breeders of thoroughbreds are expected to subscribe, yet there are many beliefs or dogmas which are firmly adhered to by the Aberdeen Angus, Galloway, and shorthorn men, as well as by fanciers and breeders of horses, and each breed may be said to have its own prophets.
Before, however, referring to a few of the articles that hold a conspicuous place in the breeder's creed, it may be well to ask the following question, viz.: Is the wild parent prepotent over the domestic? That the wild parent impresses his or her characters on the offspring more than the tame or domestic parent is the almost universal belief.
The following experiments bear on this question: - Six white rabbit does (all of which had produced pure white young to white bucks) were crossed with wild brown bucks. The forty-two half-bred offspring were at birth of a bluish-black colour (not flesh-coloured, as is the case with young white rabbits), and they soon developed a complete coat of brown hair. Now that the survivors are half- grown it is almost impossible to distinguish them from a full-blooded wild rabbit living in the same enclosure. The half-breeds are, however, not nearly so shy as the wild one, and when carefully examined they are seen to be slightly lighter in colour. Several have, as in many leverets and young wild rabbits, a few white hairs along the centre of the brow, while one has white tips to the toes of the left fore-foot. In this case the wild parent evidently prevails.
It remains to be seen whether similar results will follow when inbred does of various colours are used, and when wild does are crossed with tame bucks. It is said the white rat, when crossed with the brown, sometimes throws black rats. If this is the case the explanation may be that the white rat is an albino variety of the once common black rat. Tame albino rabbits have doubtless sprung from several breeds variously coloured. I thought some of my half-bred young would revert towards their coloured ancestors, but this, as we have seen, was not the case.
Similar results have been obtained by crossing the ordinary turtle-dove with the white Java variety. The offspring of a male turtle-dove and a white Java hen frequently, if not invariably, resemble the ordinary turtle-dove. When, however, a male white Java dove is crossed with an ordinary turtle-dove hen, a white bird is occasionally obtained. Intermediate forms never seem to occur; at the most the faintest yellow tinge is at times visible, chiefly at the tips of the wings. The cross-bred birds are very fertile inter se, and when the offspring of a white Java cock they yield about an equal number of white and turtle-coloured birds. Even when a cross-bred dark bird was mated with a pure-bred white Java bird the result was an ordinary turtle-dove, - not, as might have been expected, a white bird. With the turtle-doves as with the rabbits, the older type is prepotent. Experiments with closely inbred white birds might give somewhat different results. [* The experiments with turtle-doves were made by Mrs. Cospatrick Dunbar, partly in London and partly at Earnbank, Bridge of Earn, Perthshire.]
As already mentioned, a duck, the offspring of a black Cayuga drake and a common wild duck, produced to a wild drake seventeen young. Of these, seven take after the wild sire, the others after the black Cayuga grandsire. In this case the influence of the wild sire is not as marked as might have been expected. With the help of reversion they might very well all have assumed the dress of the common wild duck.
Hybrids between the wild pheasant and various breeds of fowls are common. Several I have examined were more like fowls than pheasants; on the other hand, a cross between a bantam cock and a grouse is described by Millais as generally more resembling a grouse than a bantam.
Of the nine zebra-horse hybrids I have bred, only two in their make and disposition take decidedly after the wild parent. As fully explained below, all the hybrids differ profoundly in the plan of their markings from the zebra, while in their ground colour they take after their respective dams or the ancestors of their dams far more than after the zebra, - the hybrid out of the yellow and white Iceland pony, e.g., instead of being light in colour, as I anticipated, is for the most part of a dark dun colour, with but indistinct stripes. The hoofs, mane, and tail of the hybrids are at the most intermediate, but this is perhaps partly owing to reversion towards the ancestors of their respective dams. In their disposition and habits they all undoubtedly agree more with the wild sire. How could it be otherwise? However hard the life of many of our horses may be, they know little of the real struggle for existence which counts for so much amongst their wild relatives. If they lack intelligence, and readily submit their backs to the burden, they are all the more thought of. In the wild state the wits are kept sharpened, and the life-saving instincts maintained at a high standard. It is doubtless for this reason the hybrids in their mental equipments are more zebras than horses; in the one case the instincts have been kept at a high state of efficiency, in the other they have been neglected. Nevertheless the wild parent is not invariably all-powerful in moulding the character of the hybrids. This is well illustrated by the three zebra-hybrids bred some years ago by Lady Meux at Theobald's Park, Hertfordshire. These hybrids are by different sires, but all out of the same zebra mare. The oldest of the three, by an English pony, is of a yellowish-brown colour, and but faintly striped. With the exception of the legs, the hog mane, and mulish tail, this hybrid in make is very like a small stoutly built cob. Moreover it is very docile and unexcitable. The second eldest is brilliantly and richly decorated with brown stripes over a bright bay background. The sire in this case was, it is said, an American pony. Again, there are zebra legs, a somewhat stout body, a plain head, and a short neck, but in disposition this hybrid is a zebra. The third hybrid is only very faintly and partially striped, and in most respects it takes after the sire, a small wall-eyed, light bay Highland or Shetland pony.
To mention yet another case. A zebra-donkey hybrid out of a zebra mare is, in many respects, far more an ass than a zebra. In this hybrid; which was bred in France, the dorsal, shoulder, and leg stripes have evidently been inherited through the donkey sire, while the head, ears, mane, and legs have as evidently the stamp of the zebra. In disposition this hybrid is far more a donkey than a zebra - no mule was ever half as stubborn. Perhaps the wild parent counts for but little in this case, because of the donkey sire having been inbred. A similar hybrid, bred in the Melbourne Zoological Gardens, though darker and less distinctly striped, is as much a donkey in its habits and instincts. It thus appears that the wild parent is not necessarily most powerful in impressing even its mental peculiarities on the offspring. This is a matter of some practical importance. It implies, e.g., that by careful selection cross-breeders will be able to produce, within certain limits, offspring differing greatly from each other, both mentally and physically. I say cross-breeders, and not hybridisers, because the difference between species and varieties is often more a difference of degree than of kind. From what has just been stated, there appears to be less difference in many ways between the zebra and the horse than between the ass and the horse, and there is about as much difference between some of the more ancient breeds of cattle and some of the newer breeds as there is between a bison (buffalo) and a shorthorn. It follows that by taking advantage of the facts already established zebra mules could be bred either closely resembling zebras or horses. By carefully selecting mares, or breeding mares for the purpose, the objectionable zebra traits might be all but eliminated without sacrificing any of the advantages such a cross may bring.
The wild parent is said to be especially prepotent because it belongs to an older and longer established type than our domestic breeds. This is very much the same thing as saying the wild parent is well-bred, or, for that matter, inbred; that prepotency amongst wild animals is, under ordinary circumstances, due to inbreeding.
Further observations will probably show that though the oldest type is likely to prevail, the offspring will take after the most inbred parent, regardless of the length of the pedigree; but when one of the parents is a sport, it is likely to transmit to at least some of its offspring its own peculiar characters, even if the other parent is inbred and wild. That sports are prepotent is supported by the results of recent experiments with insects. Dr. Standfuss, of Zurich, e.g., has, as the result of careful observation and experiment, come to the conclusion that when the normal form is crossed with a sport or aberrant variation, the result in many cases is that the offspring either take after the normal form or the sport, intermediate forms being absent.[* See paper on "Experiments in Hybridisation," by Mr. Dixey, M.A., of Wadham College, Oxford (Science Progress, April, 1898.)]
Having considered the question. Is the wild parent most prepotent? it may now be asked. Is the male prepotent over the female? This second question naturally leads to the consideration of various dogmas commonly subscribed to by breeders. It is, e.g., generally supposed that certain structures and organs are more often inherited from one parent than another. According to the teaching of the late Mr. James Howard, (1) the male parent is mainly responsible for the external structure, configuration, outward characteristics, and the locomotive system; (2) the female parent supplies the internal structure, the vital organs, and in a larger proportion than the male, the constitution, temper, and habits; (3) the purer the race of the parent the more certainty there is of its transmitting its qualities to the offspring - the parent of the purest descent having the greatest influence; and (4) apart from certain disturbing influences, the male, if of pure descent and descended from a stock of uniform colour, stamps his colour on his offspring. [* See Millais's 'Two Problems of Reproduction,' 'Our Dogs ' Publishing Company, Manchester, p. 4. ]
Even although these dogmas are widely accepted, they might have been neglected in the meantime had Sir Everett Millais not only indicated he agreed with them but in addition endeavoured to show that they were supported by the teaching of embryology. I must confess I fail to find any evidence from embryology that, as Millais puts it, "the sire influences what we can see, the colour and anatomy; and the female what we cannot see, i.e. the internal organs." Huxley, twenty years ago, said, "It is conceivable, and indeed probable, that every part of the adult contains molecules derived both from the male and the female parent, and that, regarded as a mass of molecules, the entire organism may be compared to a web of which the warp is derived from the female, and the woof from the male."[ * "Evolution," p. 29G, ' Enc. Brit.,' 1878. ] The work of embryologists since 1878 may be said to have confirmed Huxley's view. By speaking of the male as supplying the woof and the female the warp, it is not meant that the male plays the more prominent and important part, but rather that the female is especially concerned in providing the substratum on which all the organs and tissues, whether they be ex- ternal or internal, are gradually built.
When allowance is made for reversion, inbreeding, and various other factors, it is extremely difficult to estimate how far the one sex predominates over the other. As the female germ-cell (the egg) contains relatively far more extra-nuclear protoplasm (cytoplasm) than the male germ-cell, it might be argued that the female parent is more than the male concerned with the actual structure of all the organs and tissues, while the male and female take an equal part in providing the "constitution, temper, and habits." I think, however, it is safer to say, that whatever peculiarities of structure and temperament have been strongly inherited will be most readily transmitted, regardless of the sex, whether they have been recently acquired through inbreeding or derived from quite remote ancestors. Save in inbred animals, characters, even though no longer counting in the struggle for existence, which have come down from remote ancestors are likely to be strongly transmitted, whether they predominate in the male or the female; and further, characters which have during recent times, irrespective of sex, played an important part in enabling the species or variety to hold its own in the struggle for existence will also be strongly inherited. Moreover peculiarities of the nature of sports or the result of inbreeding will also, I think, be readily transmitted, quite irrespective of the sex of the individual possessing them.
It would be comparatively easy to cite instances which seem to prove that the offspring owed their colour, form, and distinctive attitudes and movements to the male; but it would be equally easy to mention cases which, seem to prove that the colour and form had been derived from the female, while the disposition, temper, and habits had been inherited from the male. Doubtless, in some cases, the male is more prepotent than the female; but in many cases the greater prepotency is the result of the male being of a purer breed than the female, i.e. of his being inbred, or because some of the characters are latent in the female. Darwin refers to the view already dealt with, that it is difficult to eradicate a blue tint when it once appears in pigeons, and also to the belief that the pouter is more prepotent than the fantail. But though blue is usually prepotent over white in pigeons, I have twice, as already mentioned, bred pure white fantails from almost blue birds, and the offspring of a white fantail cock and a blue pouter hen, though having the form and habits of the pouter, is almost white. In the last case the male was most powerful in the matter of colour, but counted for little in the form. It seems to be admitted that the male ass is more prepotent than the female when bred with the horse, - that a mule, in fact, is more like an ass than is a hinny, and that is certainly my experience; but, on the other hand, the female zebra, when bred with the horse, may prove as prepotent as the male, if the intensity of the stripes and the form of the limbs and hoofs are taken as the standard. The Dalmatian dog proved more prepotent than the sable collie, but this might have been expected from the male being, by the colouration test, one of the most, if not the most highly specialised breed of dogs in existence.

The conclusion arrived at from my experiments is, that I would not like to trust to the prepotency in the male making up for deficiencies in the female. From the two Clydesdale hybrids - full sisters - differing in so marked a manner, the one, as already explained, taking after the mare, the other after the zebra, and from members of the same family varying greatly, both mentally and physically, it seems to me breeders need not build much on the supposed prepotency of the male beyond what he has acquired through inbreeding, or by being a "sport." In all the offspring of my Arab Benazrek there is a characteristic blaze. This might be to some sufficient proof of prepotency. But a blaze may be of the nature of a "sport," and if this is not the right explanation his breeding has to be taken into consideration. He is not only well-bred, he is highly bred; whereas the dams of his offspring, with the possible exception of Mulatto, are of mixed origin, or, as we say, cross-bred. That the internal organs are not specially derived from the female I have very strong evidence. Elsewhere I shall show that a hybrid filly, out of an inbred mare and a zebra horse, is zebra-like in some of her most distinctive in- ternal organs. This is not, I think, because of the influence of the wild sire, but because these organs in the zebra are simpler than in the horse. It is doubtless easier transmitting simple ancient structures than highly specialised recently acquired structures.
From what has been already written it will be evident that I generally agree with the third and fourth articles in Mr. Howard's creed. Other things being equal, "the parent of the purest descent will have the greatest influence." But who will undertake to say what purity of descent means? It is not enough to know whether any given animal has been bred on an equal or an unequal factor system, whether one ancestor predominates or several are about equally represented. Allowance has to be made on the one hand for variation, on the other for reversion. Still, as a rule, the parent that has been longest continuously inbred to one animal is likely to be most prepotent. One might even go further than Mr. Howard as to the transmission of colour. The general ground colour in summer of zebras is not, as a rule, white. Matopo, the sire of my hybrids, is a light dun, but not nearly so light as many cream-coloured horses, the spaces between the stripes over the sides and back being of a light sand or cement colour. His less remote ancestors were doubtless still darker. In their body colour none of my hybrids take after the sire [* The light body colour in zebras has doubtless been acquired comparatively recently, hence it is never transmitted to cross-bred offspring] five take after their respective dams, four are duns, and probably take after their maternal ancestors. In their colour the zebras, with the exception of the quagga, are highly specialised. The majority of the quaggas seem to have been of a yellowish-brown colour overlaid with not very well defined stripes of a darker shade. In the Burchell and other living zebras the stripes have evidently been gradually intensified, and the ground colour lightened. Matopo has neither been able to stamp his own peculiar pattern or his own colours on his hybrid offspring. Like the Dalmatian, he seems to have bestowed the ancestral markings, but (unlike the Dalmatian) he has had less influence in settling the ground colour - this has been determined by the dams of the hybrids. The hybrids of the better bred mares are of a bay or chestnut hue - the prevailing colour of Arab foals; the hybrids of the Highland, Shetland, and Iceland mares are of a dun colour, and thus they probably take after the horses that in olden times inhabited the north temperate regions. Over 90% of the horses bred in England are, as foals, still darker than Matopo - are bays, chestnuts, or browns; and if inquiries were made it would probably be found foals are still darker the nearer we approach the Arctic circle. Of twenty Arab foals bred this year (1898) by Mr. Wilfrid Blunt, 65% are bays, 15% chestnut, 10% brown, and 10% grey, after the first coat was shed.
From the Crabbet records for the last two years it seems evident that the colour of the offspring is not necessarily determined by the sire. If one of the parents is bay the foal is likely to be bay. The bay mares (sixteen in number) had, with one exception, bay foals, although the sire in each case was chestnut: the exception was chestnut like the sire. In three cases - with a bay sire and a chestnut mare - the foals were bay, but in one case a bay sire and a brown mare produced a brown foal, while in another a bay sire and a grey mare produced a foal that became grey as the first coat was shed. It looks as if the dam had more influence than the sire, but it is really a matter of colour and inbreeding more than of sex. If one of the parents is white or grey, the other chestnut or bay, the foal is likely to be either bay or chestnut; but if the light parent is inbred the foal may (after the first coat is shed) be grey, just as it may be black if one of the parents is black and inbred. That the colour of the parent may be stamped on the offspring- even when not uniform my skewbald mare has clearly proved. In this case a uniformly coloured well-bred bay Shetland pony failed to prevent the yellow and white skewbald stamping her own unequally shaped blotches on her offspring. In Norway, as in Scotland, a considerable percentage of the foals are of a dun colour, some being from the first of a light cream-colour, others of a dark mouse-colour, but the majority are probably of a leather or red dun hue.
TELEGONY.
The belief in telegony is almost universal. That breeders and fanciers have long believed in "infection" is not to be wondered at; it is a convenient explanation of many of the obscure phenomena that come under their notice. That philosophers and literary men, social reformers and physicians, have also adopted and tenaciously hold to the "infection" hypothesis, is less comprehensible. If one may judge from the attitude of Mr. Herbert Spencer, and of certain members of the Congress of Zoology which recently met at Cambridge, and from the sayings of the literary prophet "At the Sign of the Ship," [* See Longman's Magazine, October, 1898] Weismann, and other biologists who persist in their "scepticism," may prepare for the worst. Evidently, if experiments bearing on telegony fail to prove that it occurs at least occasionally amongst mammals, it will be all the worse for science and the workers in the telegony corner of the scientific vineyard. Since I turned my attention specially to the subject in 1894 I have, in addition to making experiments likely to give telegony the best possible chance of declaring itself, investigated a large number of alleged cases of "infection." The result, so far, is that the evidence in support o£ undoubted "infection" having ever occurred is most unsatisfactory. In every case investigated the supposed infection could be accounted for by the relatively simple reversion hypothesis, and the same hypothesis I am now satisfied is sufficient to account for what at first appeared as evidence of telegony in my own experiments. I do not by any means say telegony is impossible, that it never has occurred in the past, and never will occur in the future; but I think I am justified in saying "infection" has never been experimentally produced, and that the kind of "infection" so widely believed in by breeders, if not impossible, is at least extremely improbable - as improbable as the almost equally common belief that the colour of the offspring may be influenced by "mental impressions," as Laban's sheep and cattle are said to have been influenced by the peeled wands placed before them by his son-in-law, Jacob.
What breeders understand by "infection" will be best gathered from a few examples. The race-horse Blair Athol had a very characteristic "blaze," i.e. a striking white bald face. It is said that mares, after having foals by Blair Athol, produced Blair Athol-like foals to other sires utterly unlike Blair Athol. In the same way a fox terrier, after having pups to a Dalmatian had, Millais tells us, spotted pups to a dog of her own breed. From these two cases it will be evident that breeders and fanciers believe in direct "infection." I think this kind of infection is extremely unlikely, if not impossible. My sable collie's pups to a Dalmatian have only four or five patches of colour, such as occur in foxhounds and pointers. According to the breeder's view of telegony, her next pups to a collie like herself should, if she has been "infected," be spotted like a Dalmatian. This is evidently an untenable position. Again, all my hybrids profoundly differ in the number and plan of their stripes from their Burchell zebra sire; yet, according to the popular view, the subsequent foals of their respective infected (?) dams should more or less resemble a Burchell zebra. This is an equally untenable position. The collie, if "infected," can only be expected to produce in future pups like a foxhound or pointer; the mares, if "infected," can only in future be expected to produce foals more or less like zebra-hybrids. My flea-bitten New Forest pony, after having a mule foal, has had two foals to a grey Arab with a blaze. Next year she is expected to have a foal to a bay hackney pony. Should this foal have a "blaze," before ascribing it to telegony it will be necessary to prove a blaze had not occurred for many generations in one or other of the new foal's ancestors, or was not a "sport." In the case of the fox terrier pup, before ascribing the spots to the previous Dalmatian sire it would be necessary to prove that similar spots had not occurred in some of the pup's ancestors.
Believers in telegony, with tiresome unanimity, ever revert to Lord Morton's mare. The more I consider Lord Morton's case the less satisfied I am that the mare so often referred to was infected by the quagga. A glance at Figs. 14 and 15 (pp. 66 and 67) shows that the pure-bred filly - every inch a horse in make - had far more stripes than the hybrid, and that the mane is represented as lying to one side. Darwin, in coming to the conclusion that the mare in question had been infected, seems to have been largely influenced by two beliefs: (1) that stripes are uncommon in Arabs; and (2) that the mane in the filly was upright. But extensive inquiries have convinced me that stripes are not by any means uncommon in Arabs. Mr. Wilfrid Blunt has recently been good enough to present me with a high-caste Arab foal, showing as distinct stripes in the region of the "knee" and hock as in Norwegian dun-coloured ponies; and, in addition to a dorsal band, faint indications of markings across the withers. That stripes are less common in Arabs and their thoroughbred descendants is due, I believe, to Arabians disliking duns, which they say "are only fit for Jews to ride." Were it certain the mane of the filly was, as alleged by Sir Gore Ouseley's groom, always upright, the case for "infection" would be well-nigh complete. But the evidence in support of an upright mane in the filly is far from satisfactory. The mane, as already mentioned, is represented as lying to one side in Agasse's drawing. Even in the quagga hybrid the mane was not upright, it arched to one side, as is the case, at least during winter, in all except one of my zebra-horse hybrids, and in Lady Meux's three horse-zebra hybrids.
Darwin seems to have departed somewhat from his original opinion about "infection." At least some years before his death he had come to the conclusion that telegony only rarely occurred, that it was a "very occasional phenomenon." If in the general form - the mane, tail, hoofs, and temperament - the subsequent foals of Lord Morton's mare were horses, and not at all like the quagga hybrid, the stripes are the only evidence in favour of infection. The stripes, however, if Agasse's drawings may be trusted, are the stripes of a horse rather than the stripes of a zebra or a quagga, or a quagga hybrid, i.e. they seem to have been inherited from the ancestors of Lord Morton's chestnut mare, or through the black horse with which she was subsequently mated. To make the experiment complete, she should have been mated with a horse that was not in the habit of having striped offspring, instead of with a black Arab (in which stripes were probably latent), or at least as well as with the black Arab. But even if it be admitted Lord Morton's mare was infected by the quagga, a comparison of the filly (Fig. 15) with the first sire (the quagga. Fig. 13) conclusively shows the infection was not of the kind believed in by breeders; it did not result in quagga like any more than it resulted in quagga-hybrid-like markings. At the most the quagga produced some disturbance, which caused the mare to throw back to her own ancestors.
[* It is conceivable that the previous sire, though not responsible for visible structural changes in the subsequent offspring to other sires, may profoundly influence the constitution, i.e. lead to physiological "infection." He might, e.g., confer immunity to certain diseases. ]
Breeders dread "infection" because they believe the subsequent offspring take after the first sire; that a thoroughbred mare may throw back to a cart-horse, while a mare which has bred mules is likely to throw back to a jackass. If the sire (supposing he has any influence on the subsequent progeny by other sires) leads to their resembling some former ancestor of their dam, breeders of thoroughbreds should in most cases endeavour to have their mares infected. If by putting their mares to donkeys the subsequent pure-bred foals "harked back" to ancestors some generations removed, the sooner this plan is adopted the better. Reversion to the vigorous hardy horses of bygone days would be the salvation of the English race-horse.
Though my experiments are not yet at an end, they have proved conclusively enough that "infection" does not always occur, as many seem to think. They, in fact, support the view of experienced Continental mule breeders, which is that telegony, if it occurs at all, occurs but seldom. Should it eventually be found that out of say fifty pure-bred foals, the offspring of mares that had been previously mated with my zebra stallion, there is not a single instance of "infection," the telegony dogma will be so discredited that it may in future be neglected by horse breeders; and if similar results follow experiments with dogs, rabbits, and other animals, the hypothesis ought ere long to be regarded as a mere superstition, and finally take its place with the barnacle tree and other legends of a credulous past.
In the last of the three papers. Mulatto's second foal is described at some length. Since the description was written I have seen faint stripes on the croup of two pure-bred foals, I have also seen stripes more distinct than the bands on black cats over the shoulders and sides of an old very dark mouse-dun Highland cob, and fragments of as many stripes as occurred in the quagga in a leather-dun Norwegian pony. These striped foals and horses, taken along with the foal bred by Darwin, which had a striped face as well as stripes across the croup, point to the conclusion that all the stripes on Mulatto's second foal may very well have been due to simple reversion. If next spring Mulatto, as is hoped, has a foal to a pony of her own peculiar strain, further light may be shed on this interesting but obscure problem.
Three of the four mares that in 1897 had hybrids, have each during 1898 had a foal to a horse sire; the fourth mare has again a hybrid by Matopo. It will be well to refer to these foals separately.
(a) Tundra's Subsequent Foal. - Tundra, the yellow and white (skewbald) Iceland pony, has had in all three foals. Her first, sired by an Iceland pony, was a bay; her second is the zebra-hybrid Heckla, born May 22nd, 1897; her third (born June 15th, 1898) is a skewbald, the sire being a bay Shetland pony. Heckla, as already pointed out, is very dark in colour, indistinctly striped, and in make and action like a well-bred pony. Neither in colour, make, nor disposition does the last foal show the faintest resemblance to the previous zebra sire, nor yet to her half-sister Heckla. As already mentioned, the new foal is ridiculously like her dam; the yellowish-brown patches occupy the same positions, and are of nearly the same shape and relatively of the same size, - even the mane, forelock, and tail agree in their colouring. In make and in gait, as well as in colour, the foal is her mother's own daughter, - the bay Shetland sire has apparently counted for nothing. It may be said that for two reasons evidence of "infection," even if there is such a thing, could not be looked for in the subsequent foals of the Iceland pony. In the first place she had a foal before she was put to the zebra, and in the next place she is evidently prepotent. If only the first sire is capable of "infecting," then the Iceland pony is not a suitable subject for such an experiment, but believers in telegony are not all at one on this point; in many of the alleged cases of telegony the infection is said to have been caused by the second or third mate. There can be little doubt that Tundra is inbred, at least as contrasted with many other ponies. Judging by her first foal being bay, she would not be considered inbred in Iceland, but in all probability the Iceland ponies are, on an average, more inbred than ordinary Shetland or Highland ponies. They have been long isolated on a small remote island, into which comparatively little new blood has, as far as I can learn, been recently introduced. On the other hand, had Tundra's last foal given distinct signs of infection, it would have afforded very strong evidence indeed in favour of telegony. But I may explain I selected Tundra not only to apply a severe test, but also because I wished to experiment with a mare which had already produced unstriped offspring, and because I was anxious to see whether a mare more than two thirds white would throw a lighter coloured hybrid than a black or bay mare. As already stated, the light colour of the mare counted for just as little as the light colour of the zebra; the hybrid is extremely dark, partly, doubtless, because the dam has descended from dark mouse-coloured ancestors, and partly because zebras are unable to transmit to hybrids their recently acquired light ground colour.
(b) Nora's Subsequent Foal. - Nora, as explained below, is an almost black 11.1 hands Shetland pony. Like Tundra, she had a foal before she was put to the zebra, but, unlike Tundra, she is not prepotent; instead of being a "repeater" she tends to produce offspring like the respective sires with which she is mated. Her first foal, as explained in the telegony paper, was at birth of a bay colour, and marked with a dorsal band, with bars across the legs, and a number of stripes across the withers, - was, in fact, for a time nearly as richly banded as the filly bred from Lord Morton's mare. Though a number of these stripes vanished with the foal's coat, three distinct bands persisted, as in Lord Morton's quagga hybrid, across the withers. In Nora I had evidently a mare with a tendency to produce striped offspring, but whether the first foal inherited the stripes from his dam or from his sire it was at the outset impossible to say, both parents being equally dark in colour, and, though not related, of the same strain. Nora's hybrid foal (Norette) is more of a zebra than any of my other hybrids. The body colour is, as a whole, of a leather dun shade, but in some parts it is reddish brown, the stripes are nearly black; and, instead of spots, there have been, from the first numerous well-marked stripes across the loins and croup as in the Somali zebra, and, as in Romulus, numerous rounded arches on the brow. [* From the markings over the croup of the hybrids a fairly accurate estimate may be formed of the prepotency of their respective dams.] The hoofs are longer than in the zebra sire, and the mane, though long and pendent in winter, is in summer short and quite upright. While it might be difficult for Matopo to infect the inbred skewbald Iceland pony, he had evidently an excellent opportunity of permanently stamping his characters (or those of his ancestors) on the black four-year- old Shetland pony.
Nora, having given birth to her hybrid, was put to a small Welsh pony, a bay with black points and black mane and tail. The Welsh pony being of a different breed, and having some dark hairs in the vicinity of the withers and over the croup, I at least expected Nora's third foal to have as many stripes as her first. Nora's third foal, however, now that it is five months old, as closely resembles the sire as the Iceland's third foal resembles her prepotent skewbald dam - another proof that Nora, unlike Tundra, is not inbred. The resemblance is not only in colour, but also in make and action; and, as far as one can judge, also in temperament. As in the skewbald foal, there is not yet any indication whatever, either of the previous sire, or the previous hybrid foal Heckla. Even the shoulder and other stripes present in Nora's first foal to the Shetland pony are absent. The third foal is hence less zebra-like than the first, notwithstanding the fact that the sire of the intermediate foal was a zebra far more richly decorated with stripes than Lord Morton's quagga. But though there are no stripes visible at the fifth month, there were faint indications of stripes over the croup and hind quarters at birth; while over the brow the hair, though not coloured, was so arranged that in certain lights it seemed to form a series of arches. As the first coat was shed all the indications of stripes disappeared, and I anticipate that when the winter coat is shed next spring Nora's third foal will, in her colouration, be almost identical with her sire, the bay Welsh pony.
It thus appears that in Nora we have a non-prepotent mare that tends to reproduce, not her own characters, but those of the sire with which she is mated; and yet, though extremely sensitive, there is no indication from her last foal that she had been previously mated with a zebra.
(c) Biddy's Second Foal. - Biddy, I may repeat, is a three parts bred bay Irish mare, with black points. In her colour she is more typical, i.e. she more closely approaches the dun-coloured ancestors from which horses seem to have sprung than is the case with either the skewbald Iceland or the almost black Shetland pony. Further, she is neither prepotent like the Iceland, nor yet particularly non-prepotent like the Shetland. Biddy is of about the same age, and has from the first been quite as vigorous and healthy as the zebra stallion Matopo.
In the hybrid Remus, Biddy's first foal, the stripes are dark brown, the body colour a rich somewhat light bay. Although well striped he takes after the dam about as much as after the sire, and hence is a striking contrast to Lord Morton's feebly marked quagga hybrid. In every respect I consider the bay Irish mare an excellent subject for testing the telegony hypothesis.
Biddy's second foal is by a thoroughbred chestnut horse (Tupgill) with a frontal star. As generally happens when bays and chestnuts are crossed, the foal is bay in colour. As in the dam, there are black points, and, like the sire, a star on the forehead. At birth the second foal was 35-and-a-half inches at the withers; Remus was an inch smaller - at five months 48 inches, against 44-and-three-quarters for Remus. At five months the mane was lying to both sides, in Remus the mane has from the first been upright. In the ears, tail, chestnuts, hoofs, and in every other respect, the second foal, at the age of six months, agreed with pure-bred foals, i.e. it in no way suggested either a zebra or a zebra hybrid. But during the first three months there were indications of stripes in the region of the withers, on the sides, and across the croup and hind quarters. These indications were mainly produced by a wavy condition of the hair, but in some lights several of the bands across the croup seemed to differ slightly in colour from the intermediate spaces. During the fourth month all the indications of stripes practically disappeared, and I anticipate that when the winter coat is shed next spring the second foal will be quite devoid of stripes and all other suggestions of zebras or zebra hybrids.
I may here repeat that, during the present year, I have seen similar indications of stripes over the croup of quite a number of foals. To begin with, in highly bred foals with very fine coats there are often at birth across the sides and croup, and especially in the vicinity of the flank feather, narrow markings that might be mistaken for stripes. These markings, as explained in the third paper, are caused by the hair being arranged in well-marked tracks or ridges, separated by almost hairless spaces. In these tracks, which were very distinct at birth in a cinnamon-coloured foal I bred this year, out of a bay half-Arab mare - the sire was a chestnut thoroughbred horse - we have, it may be, a restoration for a time of an ancestral condition. Sometimes, along with these hair-tracks or ridges, there are faint stripes seen only in certain lights, but evidently in part due to subtle colouring. Stripes of this nature I noticed plentifully scattered over a reddish-grey foal out of my flea-bitten New Forest pony by the grey Arab, Benazrek. More common and more evident are comparatively broad wavy bands, often seen across the croup and on the brow of half-bred bay foals. These bands may occupy the position of ancestral stripes - stripes out of which the colour has been completely washed since they ceased to count in the struggle for existence. My reason for supposing they represent ancestral stripes is based on the fact that they occupy the position of stripes in a yellow-dun Norwegian pony, and of the stripes over the croup of one of Lady Meux's hybrids, which may have been inherited either from the American trotting horse or from a remote common ancestor. I am now satisfied that foals are far more often marked with stripes - apparent or real - than is generally supposed, and that stripes will be often seen in horses if they are carefully looked for. From this it follows that the stripes on the "colts" bred by Sir Gore Ouseley out of Lord Morton's mare by the black Arab horse are, after all, not so very remarkable. The zebra, Matopo, having failed to "infect" four mares, it is quite evident that telegony does not invariably occur, as many breeders believe. It remains to be seen whether it occurs in even 1 or 2 per cent, of cases, as was supposed by Romanes. I have now eight mares that, if telegony is true, may have been infected. I hope to continue breeding from these mares and from their offspring for some years to come, in order to give the supposed infection an opportunity of showing itself. If, out of fifty or a hundred of the descendants of these mares, a single case of undoubted infection is obtained, the fact of telegony will be established; if, on the other hand, not one out of a hundred foals "harks back" "infection," if it occurs at all, may be treated as a negligible quantity.
In addition to the experiments with mares, I have been on the outlook for evidence in favour of telegony in rabbits and dogs, fowls and pigeons, I first made sure that six white rabbit does produced white young to white bucks. The white does were then crossed with wild bucks, with the result, as already stated, that they brought forth over forty young that could with difficulty be distinguished from wild rabbits. When next bred with a white buck, they all produced perfectly white young, proving, as far as colour could show, that none of the does had been infected. I propose making further experiments with rabbits next summer.
The results obtained with fowls and pigeons also failed to give any support to the telegony hypothesis. The experiments with dogs already completed, have not supported the deep-rooted and wide-spread belief that, in dogs, at least one of the subsequent offspring almost invariably takes after the first (or previous) sire if it happens to be a mongrel, or to belong to a different breed from that of the dam.
SATURATION.
Almost identical with telegony is the saturation hypothesis. Bruce Lowe, the prophet, if not the originator of the saturation theory, defines it as follows: - "Briefly put, it means that with each mating and bearing the dam absorbs some of the nature or actual circulation of the yet unborn foal, until she eventually becomes saturated with the sire's nature or blood, as the case may be." [* Bruce Lowe, 'Breeding on the Figure System.' ] That something of this kind occurs in the human family is also widely believed. Bruce Lowe, I understand, meant to account for telegony by the saturation theory, but evidently the saturation theory goes further than telegony; it seems to imply an actual change in the organs and tissues of the dam, - she is said to be "saturated with the sire's nature or blood;" whereas, according to Weismann, Romanes, and others, telegony, if it occurs, only influences the germ-cells, which stand in much the same relation to the parent that a cargo of wheat does to the ship conveying it. Evidence of saturation is looked for, not so much in a change in the appearance of the female parent, though a change of this kind is sometimes insisted on, as in the offspring. They are said to become progressively more and more like the sire, the second foal taking more after the sire than the first, the third than the second, and so on. Now, as it happens, a paper was read at the recent Zoological Congress at Cambridge by Professor Hubrecht of Utrecht, which seems to lend support to this theory as well as to the telegony theory. The object of this paper was to show that blood-corpuscles in some mammals pass bodily from the foetus into the tissues of the dam. It is not asserted that this happens in the mare; on the contrary, the inference is that in the horse, pig, and certain other families this passage of the foetal blood-corpuscles does not occur. But even if it be admitted that the "actual circulation" or blood of the unborn foal is absorbed, it does not necessarily follow that the nature of the dam is in any case gradually altered, except in as far as the foetal corpuscles are the bearers of the germs of disease. The saturation theory may also be said to be supported by the recent investigations of M. Charrin, who found that "diphtheritic toxins injected into the embryos of a pregnant rabbit caused the death of the mother on the fifth day;" [* The Medical Press, October 5tli, 1898, p. 360.] and further, that a rabbit could be rendered immune by injecting protective toxins into the embryos. M. Charrin infers from his experiments that the characteristics acquired by the mother from one set of embryos could be handed on to other embryos by a different sire. But even should further experiments justify this conclusion, it does not follow that there is either telegony or saturation." [* Experiments with toxins might prove that physiological "infection" occurs.]
I have been led to refer to saturation because the second hybrid of the Clydesdale mare is more a zebra than her first. Some seem to think the Clydesdale has already been "saturated" by the zebra. It is a matter of common knowledge that the members of any given family, litter, or brood by the same parents vary considerably in colouration, form, and disposition. That the second Clydesdale hybrid differs from the first is hence not a unique, nor yet an unexpected phenomenon. But though the members of the same family often differ from each other, it by no means follows that the cause of this variation has yet been discovered.
If the dam in form, &c., gradually assumed the characters of the sire, and if the resemblance of the offspring to the sire proceeded at a progressive rate, there would be a presumption in favour of saturation. But undoubtedly this is not invariably the case, and when it does happen that the female appears to assume the characters of her mate, or the offspring to become more and more like their sire, a simpler, or at least a more scientific explanation will doubtless be available. In large families the youngest children often take more after the mother than the eldest. Of Lady Meux's three hybrids out of the same zebra mare, the youngest is undoubtedly most like a horse, but the intermediate one is by a long way most like a zebra. The Clydesdale's second hybrid is far more a zebra than her first, not in colour, but in make and gait and disposition. If the striking difference between the two full sisters is not due to saturation, how can it be explained? A possible explanation will be easier if we consider for a moment some of the results of cross-fertilisation. It is almost inconceivable that two germ-cells derived from two separate animals or plants can ever be identical either in form or structure. Hence, when two germ-cells - cue the representative of a male, the other of a female animal or plant, more faithful representatives than any two members of Parliament can ever hope to be of their respective constituents, - when these somewhat differently constituted germ-cells meet and completely fuse, the new cell formed will (except under special and unusual circumstances) differ not only from each of the two original cells, but probably also from every cell that has ever previously existed or will exist. If an attempt were made to sort the members of two large electoral districts, it would be found impossible to divide them into couples that in every respect agreed with each other; the difference would of course be greater if the females of one side were ranged with the males of the other. Differences in form and colour would be at once evident, but the differences in temperament might be even greater. If the districts happened to be far apart (one in, say, the south of Ireland, the other in the north) all the differences would probably be accentuated. The units of protoplasm making up the essential parts (the chromosomes) of two germ-cells doubtless often differ quite as much as the members of two constituencies - differ in form as well as in affinities. To make the comparison at all complete between two germ-cells and the electors of two districts, it would be necessary to include not only the women and children, but also the less remote ancestors, and at least the ghosts of the remote ancestors. If in such a collection only those who could find mates the image of themselves in structure and temperament were allowed to live, long would be the list of the doomed. Among the survivors the less remote ancestors would be well represented, while amongst the doomed the representatives of quite recent generations would be extremely abundant. The reason of this being that in bygone times our islands were inhabited by perhaps a single nearly uniform race, whereas, during recent centuries, wave after wave of new types has reached our shores, great waves of Celts, Romans, Angles, and Norse, and smaller waves of Slavs, &c. In the same way, if only the units of the two germ-cells which agree in structure and have a certain affinity for each other are capable of forming stable combinations, there must often be millions of units that fail to enter into the composition of the new cell.
According to Mr. Galton's law, the immediate parents, on an average, only actually provide half of the units composing the new cell, the other half being provided indirectly by the ancestors. But evidently the extent to which the immediate ancestors are represented in the new cell (out of which the new individual is eventually formed) will, other things being equal, depend on how much the individuals (parents) from which the cells came essentially agree with each other. If alike and closely related the immediate parents might be represented by more than half the units; if unlike and unrelated, by considerably less than half the units in the new cell - the potential new individual. If one parent happened to be prepotent and the other not, only one of the parents might predominate in the new individual.
From what has been said it will be evident that quite a number of results may follow the union of two germ- cells. Under certain conditions a considerable departure may be made in one or more directions, and lead to the appearance of something strikingly new, i.e. to the appearance of a sport. Instead of marked progress, there may be marked retrogression towards a remote ancestor. This when the units repel rather than attract each other, or unite to form unstable and short-lived combinations. As a rule, however, the result is neither a marked step in a new direction nor a long step in a backward direc- tion. There is often slight progress in one or more directions, i.e. normal or continuous variation, accom- panied by slight regression in others.
Instances of regression we have (1) in the case of the last Iceland foal, which is an exact reproduction of the mother; [* This might be either considered an example of prepotency or regression to the mother's ancestors.] (2) in the Angora rabbit, which is a restoration of the grandmother; (3) in the collie-Dalmatian pups, which are apparently a reappearance of fairly remote ancestors; and (4) in the fantail-owl-archangel pigeon, in which we have an almost complete reversion to the common ancestor of all the pigeons. Examples of progressive and of a combination of progressive and regressive changes might also be given.
To return to the Clydesdale's second hybrid. Is it more a zebra than the first because the mare has been saturated by her zebra mate, or can the difference be otherwise accounted for? I think in the second hybrid we have evidence of further reversion than in the first to the ancestors of the dam, and at the same time more of the characteristics of the zebra or o£ the ancestors of the zebra. The Clydesdale's second hybrid (Black Agnes) is the darkest of all the zebra hybrids I have bred. I have already stated that mares have more influence in determining the ground colour than the zebra, probably because they have not departed so far as the zebra from the original colouring. Unfortunately I have been unable to find out the colour of the recent ancestors of the Clydesdale mare; but as the white fantail pigeons resemble a white grandparent instead of their blue parents. Black Agnes probably takes after a grandparent having latent stripes. This would not only account for her being a nearly black-mouse dun, but also for the faint horse-like stripes across the croup.
The numerous stripes proclaim the influence of the zebra sire, but quite as suggestive of the zebra are the make, movements, general habits, and intense vigour, in all of which Black Agnes is a striking contrast to Brenda. From the outset the second hybrid has been more intense and more vigorous, and more restless when away from her dam, than Brenda. It is conceivable that the dark colour of Black Agnes is partly due to the fact that she is a stronger and more vigorous foal than her full sister Brenda, for intense energy often expresses itself in deeper or richer colouration.
Taking all the facts into consideration, instead of ascribing the difference between the first and second Clydesdale hybrids to saturation, I am inclined to ascribe it partly to more pronounced reversion, but mainly to better nutrition - to a greater ripeness - of the germ-cells from which it originated.
In the same way I account for the difference between members of the same family, and between full brothers and sisters of the same litter or brood. Sometimes the difference is due to abnormal or discontinuous variation - to some of them being sports, - sometimes to more or less marked reversion to the ancestors of either parent, but in the majority of cases I believe it is mainly a question of nutrition. [* As an instance of the influence of nutrition during development, I may mention that I recently found that eight rabbits in one uterine horn were exactly the same weight as four rabbits in the other horn. This looked like a case of superfoetation, but as the eight small foetuses were as well developed as the four large ones, the difference was evidently due to the same amount of nourishment being provided for each horn regardless of the contents.]
The experiments with Echinoderms already mentioned especially suggest this. [* Vernon, Roy. Soc. Proceed., May 26th, 1898.] Whether the hybrid Echinoderms resembled the one species or the other depended on the relative degree of ripeness of the germ-cells. At the beginning of the season, when the germ-cells of the one species (Species A) were unripe, the hybrids resembled the other species (Species B); but as ripe germ-cells of Species A increased, the hybrids became more and more like A. It is equivalent to saying the prepotency of the germ-cell, other things being equal, varies with its ripeness, or, in other words, its state of nutrition. Abundant evidence could be given to prove that there is, ns a rule, an intimate relation between the nutrition of the germ-cells and the nutrition of the individual in which they are stored. Just as the grains of wheat may materially suffer from the bad state of repair of the ship in which they are carried, so may germ-cells suffer when the body in which they are lodged is out of condition, ill-nurtured or over-strained. As the grains of wheat too early harvested may produce an indifferent crop, so may immature germs produce indifferent offspring. To summarise, I believe experiments may show that the difference between members of the same family, and members of the same brood or litter, is mainly due to the structure and condition of the germ- cells, and to the nature of the combinations formed when the carriers of heredity, the protoplasmic units derived from two separate individuals (the parents), actually combine to form the new cell out of which the new individual is gradually developed.
STERILITY IN EQUINE HYBRIDS.
It is not in obedience to any natural law that hybrids are, as a rule, sterile. Formerly zoologists were ever ready to consider intersterility as strong evidence of specific distinction, even when the mutually sterile forms were almost identical in structure and habits. Were structure alone taken as the standard, not a few of: the animals that are looked upon as mere varieties would be raised to the rank of species, while a number of species would be degraded to the rank of varieties. Even as it is, a number of species yield when crossed fertile offspring. For example, hybrids between the common goose and the Chinese goose - two very distinct species - are perfectly fertile, as are hybrids between the common duck and the pintail duck, and between various species of pheasants. The Indian buffalo and the American bison produce fertile hybrids with the wild ox of Europe; while the common humped cattle of India yield fertile hybrids with the domestic ox.
Amongst plants, hybrids are sometimes quite fertile; while some crosses are quite, or almost, sterile. There is no hard and fast line between species and varieties, and hence there can be no fundamental difference between a hybrid and a cross, nor yet any a jjriori reason why any given hybrid should be sterile, or any given cross fertile.
It is no longer possible to contend that species were originally endowed with mutual sterility by way of preventing the confusion that would result from free interbreeding. Sterility has doubtless been acquired in some cases slowly, in others abruptly, but how it has been acquired it is impossible in most cases even to guess. Intersterility has not hitherto been acquired by any of our breeds of pigeons, or dogs, or horses, or cattle; but this may be due to their comparative youth, or to the artificial conditions under which they were produced and live. Romanes believed that the moment any variety became sterile, or nearly sterile, with other varieties and with the parent species, that same moment it mounted the first rung of the ladder leading to specific rank. That sterility between varieties would improve their chances of developing into species is sufficiently evident; but that in the majority of cases sterility, however acquired, is the starting-point - the password - towards specific distinction has not been established.
Mr. Galton thinks prepotency is a highly heritable sport. Sterility may also be a sport, an aberrant variation, not due to or associated with any demonstrable structural changes.
It seems to me experiments might show that preferential mating accounts for much. Horses are quite remarkable for their strong likes and dislikes, and the same may be true of many other animals. Preferential mating (apart from sterility) might, through inbreeding, gradually build up distinct more or less intersterile prepotent varieties.
That intersterility may occur without any apparent change in the reproductive organs is extremely probable. This is suggested by the fact that a large number of wild animals invariably fail to breed - are virtually sterile - in confinement. But intersterility seems to arise without even any appreciable change in the environment, and it is well known that a not inconsiderable number of animals, though perfectly formed, are quite sterile; or, what is even more suggestive, sterile with some individuals but not with others, or sterile until the system receives some sudden shock, or until changes are gradually produced by a different kind of food. In discussing sterility, it is generally pointed out that of all the systems the reproductive system is most sensitive to changes in the surroundings. If the reproductive system is especially sensitive, if, e.g., by inappreciable changes in the surroundings fertility is increased, impaired, or completely lost, need we be surprised that hybrids (in which the reproductive system is of necessity modified by regressive and other changes) are frequently sterile?
But it will be asked. What evidence is there that the reproductive system of hybrids in any way differs from that of the parent species? Strange as it may seem, there does not yet appear to exist an account of the reproductive system of either mules or hinnies, or, as far as I can discover, of any mammalian hybrids. Female mules may, as is now and then alleged, be fertile, while male mules may be invariably sterile, but until the germ-forming glands of both male and female mules and hinnies have been systematically investigated it is hardly safe to hazard an opinion on the subject in the absence of well-authenticated cases of fertility. I have not yet had an opportunity of examining either the germ-forming glands of ordinary mules or hinnies, but I have recently had the chance of learning something of the reproductive glands of zebra hybrids.
Sterility of Male Zebra Hybrids. - In the two-year-old hybrid, Romulus, the reproductive apparatus seems to be fully developed, as are the reproductive instincts. As in his zebra sire, there are large teats, but, unlike both sire and dam, though doubtless like the ancestors of both, the skin forming the scrotal sac has a complete coating of hair; this is doubtless another instance of regression.
But notwithstanding the fact that Romulus is apparently as well developed as a horse of the same age, he has not yet reached maturity, i.e. his germ-cells are still incompletely formed; there is a head, but the merest rudiment of a tail. This may, however, be accounted for by his mixed origin. Though yearling colts and fillies have been known to breed, zebras never seem to reach maturity until the fifth or sixth year. Hence, though zebra hybrids are not fertile as two-year-olds, they may prove fertile when three or four years old. If one may judge by the behaviour of the male germ-cells in other mammals, it is possible that a change of the surroundings - of the food, temperature, &c. - might make a profound difference, transform a sterile hybrid into a fertile one.
Domestication is known to hurry on maturity. In a wild state it would be a disadvantage for a young stallion to reach maturity much before his full strength is required, until be has some chance of successfully challenging the old leader of his troop, or forming for himself a new one. It may safely be taken for granted that the wild ancestors of our horses were later in reaching maturity than their pampered domesticated descendants. Hence, apart from any retarding influence the zebra may have had, Romulus, in as far as he has regressed towards his maternal ancestors, will presumably be late in reaching maturity.
In addition to Romulus, I have for some time had a male zebra-ass hybrid under observation. This hybrid having unfortunately died, I shall elsewhere be able to describe at length the condition of the germ glands. In the meantime I may, however, say that, as in Romulus, the reproductive system of the three-year-old zebra-ass hybrid seemed quite perfect, but, as in Romulus, the germ-cells were incompletely developed, and hardly at all mobile, owing to the tail being only on an average twice the length of the head; in the horse and zebra the tail is quite eight times the length of the head. This condition of the male germ-cells is better described as arrested development than as regression; they closely resemble a phase in the development of the corresponding cells in the horse, but do not necessarily reproduce the mature germ-cells in any of the ancestors of the Equidae. Whether, had this hybrid survived, ripe germ-cells would have eventually appeared I am unable to say, and I am also unable to say whether a different diet or different surroundings would have resulted in the formation of perfect germ-cells.
I had some hope that this hybrid would be fertile, for in its breeding it agreed with a hybrid (referred to by Darwin and others) which is said to have had offspring when mated with a pony. In the 'Gleanings from the Knowsley Menagerie ' there is a figure (lix, 2) of this triple hybrid, and at p. 73 it is described as follows:- "The offspring of a mule (the produce of a male ass and a zebra) with a bay mare pony. Iron-grey with a short narrow cross-band on withers; very faint indications of stripes on the sides, and more distinct dark stripes on outside of the hocks and knees; tail bushy from the base like a horse, head heavy, mane brown and grey. This animal used to draw a small cart. It stands eight hands high." Darwin, writing in the seventies, says of this cross, "Many years ago I saw in the Zoological Gardens a curious triple hybrid from a bay mare by a hybrid from a male ass and a female zebra."
Not a few mistakes have been made about hybrids. There is, for example, the famous case of the fictitious cross between the hare and the rabbit, and one occasionally hears of a hybrid, equally improbable, between a dog and a fox. Yet it is not likely Darwin would make so pointed a reference to this triple hybrid had he not satisfied himself that it was genuine. An effort is being made in the gardens of the Melbourne Zoological Society to induce a zebra-ass hybrid to breed. This hybrid is certainly now old enough, and, with its genial climate, Melbourne seems peculiarly well adapted for an experiment of this kind. It need hardly be urged that the first possible opportunity that offers should be taken to examine the male germ-cells of ordinary mules and hinnies. This could be best done where mules are common, and where specimens of various ages are available.
Sterility of Female Zebra Hybrids. - Periodically it is asserted in the Field, or in some other newspaper, that a female mule has produced a foal. Sometimes the interesting event is said to have taken place in the East, sometimes in the West. Why it should always occur in some out-of-the-way district is hard to explain. The last account of a female mule having a foal comes from India. The writer, Veterinary Captain W. D. Gunn, seems to have satisfied himself that the mule is the real mother of the foal in question, and not merely a foster-mother. Mr. Gunn being a trusted and able member of his profession, we may perhaps assume that he has come across an undoubted instance of a fertile mule. Unfortunately, though he may have satisfied himself, he can hardly be said to have satisfied the readers of the Field. But it will be well to allow Mr. Gunn to speak for himself; he says, "A most unusual event occurred in the Kapurthala State, India; indeed, it seems to be the only case on record. A mule, belonging to a potter in the above state, gave birth to a male foal on the day after its return from the Tirah Field Force. Parturition occurred on August 6th during the night, and on information being given to the Prime Minister of the state, Sirdar Bhagat Singh, C.I.E., he at once went to see it early the following morning. The greatest excitement has been caused in the town of Kapurthala by this extraordinary occurrence, and the pundits are all at a loss to know what to think about it. They say that such an event has never been known before, and the Hindu shastras say that whenever a mule becomes pregnant it must die before giving birth to the young. Large crowds go daily to see the mother and foal, and the pundits are consulting the stars and shastras as to what is portended by the event.
"When Sirdar Bhagat Singh saw that the mule had really dropped a foal he at once communicated with the Civil Veterinary Department, find after making further inquiries I proceeded to the Kapurthala State and took the photographs which I send you by this post. Veterinary Captain Joslen, officiating principal of the Lahore Veterinary College, accompanied, and he will vouch for the correctness of these statements. The mule must have been covered by a pony while proceeding with the transport to the frontier war. As will be seen, the foal is beautifully formed, and has the appearance of a pony foal with very small ears. The mother is about twelve years old, and stands 11-and-three-quarter hands, and is a very typical Indian transport mule."
After referring to a number of doubtful cases of fertile mules, he adds: "In the present case, however, there can be no doubt about the genuineness. The foal was dropped at midnight, and was seen the next morning by large crowds, including the Prime Minister, a Sikh gentleman of the highest respectability."[ * The Field, September 17th, 1898]
Every breeder knows that mules and young virgin mares occasionally give milk, and also that foals are at times enticed away from their mothers. Mr. Gunn may have assured himself that the "beautifully formed" foal with "very small ears" has not been stolen by the mule in question from a pony mare. Unfortunately this possibility is not referred to in his letter. Although my experience is but limited, I have had an instance of a foal being stolen by one mare from another, and also an instance of a mare leaving her foal to take care of itself immediately after it was born. Mr. Gunn, or his companion Veterinary Captain Joslen, of the Lahore Veterinary College, may be able to clear away all doubts; and better still, the mule and foal may ere this have found their way to one of the Government stud farms, where steps will be taken to give the mule a chance of again breeding, and where later the fertility of the foal will be tested.
Mr. Tegetmeier having long taken an interest in the subject under consideration, it may be well to indicate what he said on the subject about a year ago. In the Field of October 23rd, 1897, he writes, "I have long held the opinion that the ordinary equine mule of both sexes is sterile. This, as is stated in the work on 'Horses and Mule Breeding' by Mr. Sutherland and myself, has been founded on the great experience of M. Ayrault, and the belief has been traced by us to the phenomena of induced lactation in the mule mares, many of which have been known to suckle offspring not their own. At the same time I am free to acknowledge that many cases of strong evidence in favour of the fertility of mule mares have come before us." After referring to a case he had just heard of in Mexico, he proceeds to say, "This is one of the most detailed accounts of a fertile mule that has come under my notice." Mr. Tegetmeier, it will be observed, does not say what he actually thought as to the genuineness of any of the cases he had recently heard of, but it may be inferred that in 1897 he was less sure mules were invariably sterile than he was when the work 'Horses, Asses, and Zebras' was published in 1895. As "little scientific observation has been brought to bear upon the question of the character of the hybrids between the horse and the ass," I shall now state shortly the results of recent observations on the reproductive system of two female zebra mules, or, to be more accurate, of a female zebra mule (zebrule) and a female zebra hinny (zebrinny).
In my zebra-horse yearling hybrid, Heckla, which died recently, the reproductive organs were found to be almost identical with the corresponding organs in a young Burchell zebra (which died about a year ago), and unlike in various respects those of a mare. Heckla thus inherited her reproductive system through her sire, not from her dam, the Iceland pony. In a ten-year-old horse-zebra hybrid which also recently died I found the reproductive system resembled that of Heckla, but there was this important difference, that one of the reproductive glands contained one large and several small Graafian follicles which closely agreed with the follicles in the reproductive gland in the mare, and were, as far as I can make out, identical with the germ sacs or follicles in a six-year-old Burchell zebra. One of these follicles had a diameter of 1-and-a-quarter inches, - a ripe follicle in a sixteen-hands mare has a diameter of about 1-and-three-quarter inches. From the ap-pearance of the follicle it may very well have contained a nearly mature germ-cell (ovum).
While it may be safely inferred male germ-cells almost devoid of flagella or tails are not likely to be of any use, it is practically impossible to say whether any given fermale germ-cell would, if fertilised, develop into a new individual. The fact, however, that in the oldest of the two zebra hybrids examined there was a follicle enclosing presumably a nearly ripe germ-cell favours the view often advocated, that female mules are at least occasionally fertile, and at the same time encourages us to believe that the Kapurthala potter's mule is the actual mother of the foal found in her possession last August. I ought, however, to say that a three-year-old mule (ass male-horse female hybrid) from the New Forest has so far proved sterile alike with Shetland and Welsh ponies, and with the zebra female-ass male hybrid already mentioned, and that a nine-year-old zebrinny (horse-zebra hybrid) seems also to be sterile with both Arab and Clydesdale horses.
I intended discussing in this introductory chapter the habits and instincts of young foals, the rate of growth of foals and hybrids, and the use of stripes in the Equidae; but a consideration of these and other questions may very well be deferred until I have completed the first part of a memoir 'On the Development of the Horse,' and a small work 'On Zebras and Zebra Hybrids.'
I have, in conclusion, much pleasure in expressing my great indebtedness to Lord Arthur Cecil, Mr. Wilfrid Scawen Blunt, the Hon. Walter Rothschild, and Lady Meux. I am also not a little indebted to Sir William Flower, K.C.B., F.R.S., Mr. Oldfield Thomas and Mr. Pocock of the Natural History Museum, London, Professor Mettam, B.Sc, of the Royal (Dick) Veterinary College, Edinburgh, Mr. Dixey, M.A., of Wadham College, Ox- ford, and Professor Stewart, F.R.S., of the Royal College of Surgeons' Museum, London. To Mr. Alfred E. Pease, M.P., Mr. Rowland Ward, Mr. Arthur H. Neumann, and Mr. C. V. A, Peel I owe my best thanks for the loan of valuable skins; and I must add that I owe much to the writings of Mr. Darwin, Mr. Tegetmeier, Sir Walter Gilbey, Bart., and Herr von Nathussius.

Fig. 1. Matopo.
Fig 2 - Mulatto
PART I.
A - THE BIRTH OF A HYBRID BETWEEN A BURCHELL'S ZEBRA AND A MARE.
THE BIRTH OF A HYBRID.
In a note on Telegony, [* Appendix] published in 1895 in The Veterinarian, I suggested a number of experiments which might be made with a view to settling, if possible, whether a previous sire has any influence on subsequent progeny obtained by other sires. At the same time I mentioned that I was making arrangements to carry out three of the experiments suggested, viz. (1) To cross a number of mares with a male Burchell's zebra, and then mate them with an Arab or other suitable horse. (2) To cross a zebra mare with an Arab horse, and then mate her with a zebra. (3) To cross an ass (Equus asinus) with a zebra horse, and then mate her with a jackass.
Having begun in 1894 to make arrangements for the telegony and other investigations, I was in a position early in 1895 to begin operations, - that is to say, I had secured a large area of grass land and a small paddock, provided accommodation for and purchased three Burchell's zebras (a horse and two mares), an Arab horse, and a number of mares, including a thoroughbred filly by "Petrarch," a mare by "Gunboat,'" and mares from Ireland, Iceland, Shetland, and Norway. Further, Mr. Wilfrid S. Blunt, on hearing of the proposed experiments through the late Professor Groodhart, was generous enough to send me an Arab mare ("Bernabit"); and Lord Arthur Cecil, whom I had the good fortune to interest in my work, lent me "Mulatto," one of his Island of Rum ponies, and later added to my stud a young New Forest donkey.
It is hardly necessary to point out that even should the experiments in hand settle, or at least throw new light on the question of telegony, final results are not likely to be gained until several years have elapsed. However, as the investigations are not limited to testing the influence of a previous sire, but either directly or indirectly deal with the development and ancestral history of the horse, with reversion or atavism, and polydactylism, and with various other problems of a more or less interesting nature, it may be well to put on record now some of the events that have happened since the work was started.
The Birth of a Zebra Hybrid. - The most interesting event I have to chronicle is the arrival of a hybrid between my Burchell zebra stallion (Matopo) and the West Highland pony (Mulatto). The hybrid, which, as announced in the Field [* The Field, August 22nd, 1896], was born on the 12th of August, appears to be the first cross obtained between an ordinary mare and a Burchell's zebra stallion. Partly on this account, and partly because zebra hybrids may help to solve the transport difficulties in various parts of Africa, and do something towards overcoming the aversion for mules that has so long prevailed in England, a short account of the new arrival may not be unacceptable. But before speaking of the hybrid colt I ought to say a few words as to his parents. There are still three zebras found wild in Africa - two mountain zebras (E. zebra and E. grevyi) and the zebra of the plains (E. burchelli). The sire (Fig. 1) of the hybrid, which belongs to the Burchell group of zebras, I obtained from the Antwerp Zoological Gardens.
Mr. G. R. de Courcy-Perry, H.B.M.'s Consul-General at Antwerp, to whom I am indebted for timely help at a critical moment, reported that the directors of the gardens considered their zebra stallion a superb animal; he is certainly the most handsome specimen I have ever seen. While in Antwerp "Matopo" proved a successful sire, but it was only found possible to mate him with zebras of his own species. However, as a zebra long expected from South Africa never arrived, and as the Antwerp one was the only available mature male Burchell's zebra in Europe at the time, I had either to secure him or give up the experiments for another year. Matopo, last summer (1895), was certainly very fastidious; instead of a large crop of foals I am only able to report the arrival of one; but as a cross between a male Burchell's zebra and a mare has apparently not been hitherto obtained, I ought perhaps to be thankful that the telegony experiments did not prove a complete failure. By carefully studying the habits of the zebra mares and Matopo all the difficulties have been practically overcome; he no longer reminds one of the proverb, "You may lead a horse to the water, but you cannot make him drink." I need only add that the zebra horse, though low at the withers, stands nearly thirteen hands high; that he is beautifully marked, and in his form and movements suggests a horse far more than either of the mountain zebras; further, his action when trotting is even more perfect than that of his stable companion the high-class Arab horse Benazrek.
Mulatto (Fig. 2), the dam of Romulus, is a West Highland pony, thirteen hands (52 inches) high. Lord Arthur Cecil, who has taken a lively interest in the investigations from the outset, first intended sending a couple of New Forest ponies. After further consideration he, for various reasons, selected Mulatto; this has proved a fortunate selection. Apart from the all-important fact that Mulatto has produced a foal to the zebra, she is in many ways preminently suitable for the experiments in hand. From information kindly supplied by his lordship, it appears that the breed to which her sire belongs has been for many years all but completely isolated on the Island of Rum, a small island lying between the mainland of Scotland and the Outer Hebrides. As far as is known, fresh blood has only once been introduced during recent times into the Island of Rum. This was in 1848, when the then proprietor of the island, the late Marquis of Salisbury, sent to Rum a stallion belonging, it is believed, to the characteristic West Highland strain. Mulatto's dam came from the Long Island (Outer Hebrides), but she belonged to the same breed as the sire. Like Mulatto and all her ancestors, as far as they can be traced, the dam was almost black, and, like the majority of this particular breed of ponies, her eyes were of a hazel colour - not brown, as in the majority of horses. In Mulatto there is only a faint indication of the characteristic hazel-coloured iris. It is difficult to account for the existence of more or less isolated troops of well-bred ponies in the Western Highlands. The late Marquis of Salisbury believed that, notwithstanding their colour, they had Eastern blood in their veins. It has been suggested that they numbered amongst their ancestors horses which escaped from the ill-fated ships of the Spanish Armada. In support of this belief it may be mentioned that an old tapestry in the House of Lords (a representation of which appeared some years ago in the Illustrated London News) indicates that storms overtook the Spanish fleet at several points off the Western Islands. It may be more than a coincidence that well-bred ponies were afterwards found on islands adjacent to the storm areas - on islands and parts of islands to which the dismantled Armada ships might very well have drifted. Further, the Mulatto breed of ponies resembles well-bred black horses often met with in Spain at the present day. Whatever the origin of the ponies in question, it is enough that they belong to a distinct breed, and that probably only once (in 1848) during many generations has fresh blood been introduced into the isolated and somewhat inaccessible Island of Rum, from whence Mulatto's sire was exported in 1888. As a proof of the isolation, or, in other words, of in-breeding, crosses between Island of Rum ponies and other breeds present nearly all the characters of the West Highland race.

Fig. 3. - Romulus (seven days old) and his dam, Mulatto.
Fig. 4. - Romulus, twenty-seven days old.
Mules generally strongly resemble their asinine progenitors, but the mules bred by the late Marquis of Salisbury in the Island of Rum were observed on reaching Hatfield to resemble ponies rather than donkeys. This, to my mind, proves that Mulatto belongs to a well-marked and distinct breed, and that whatever the result may eventually be, she is in every way as suitable, as well adapted for taking the place of the nearly purely bred Arab mare used in Lord Morton's famous experiment, as the zebra Matopo is an excellent substitute for the now extinct or all but extinct quagga. It is only necessary to add, before proceeding to describe the hybrid, that the foals of the black Highland ponies are frequently at first mouse-coloured, with in some instances a faint dorsal stripe, and a patch of dark hair at the shoulders, which represents the bands so often seen in dun-coloured Norwegian ponies. The dorsal stripe usually disappears soon after birth, and the mouse-colour never reappears when the first or foal's coat is once shed.
Turning now to the hybrid, I may first mention that the period of gestation was normal, 342 days. With the mare it usually varies from 340 to 350 days. In the zebra, as in the ass, it is said to extend over twelve months. Within a minute after birth the hybrid (Fig. 3) was rushing about as if he were a young zebra, whose existence depended on his at once joining the troop of which his dam was a member. Being extremely alert, and ready to gallop off at any moment, he seemed at a loss to understand the inaction of his placid dam. Though when nearly a month old (Fig. 4) Romulus was no longer so restless, or like a timid young animal whose only safety was in flight, he was still surprisingly energetic, and he seemed to enjoy nothing better than coursing about his paddock as if he were escaping from some dreaded foe. He is now wonderfully tame, and courts rather than shuns notice; and from the first he has behaved himself quite differently from a young New Forest mule I had the opportunity of constantly watching during last summer. In nearly all his movements Romulus resembles his sire rather than his dam; he has not the dainty action of a young mule or a young donkey, and yet he differs in his gait from a horse. As he grows older I anticipate the beautiful action of the zebra will become more and more apparent. The Arab mare, Bernabit, when set free in the field carries her head high in the air in a most suggestive way as she gallops about. The zebra horse, on the other hand, carries his head low, and twists his neck as if engaged in single combat, ready to seize the limbs of an adversary. In his gambols the hybrid carries himself like a zebra, and this without once having had an opportunity of seeing Matopo disporting himself.
When in the field with mares the zebra horse herds them most jealously; when anyone appears on the scene he gallops along uttering his war-cry, prepared to defend his troop against all comers. I shall not be surprised should Romulus imitate his sire in this also, should an opportunity offer by-and-by.
As the time drew near for the birth of the hybrid I became more and more curious as to which of its parents it would most resemble in shape and colour. When two distinct types are crossed the progeny may present the characters of both parents, or may closely resemble one of the parents, or by reverting towards the ancestral type differ decidedly from the immediate ancestors. For example, mules sometimes closely resemble donkeys, at other times they resemble ponies. Again, when a fantail is crossed with a pouter, the young sometimes resemble a blue rock, the supposed remote ancestor of all the pigeons. There has probably been no intercrossing for many thousands of years between zebras and the other wild Equidae, and it is quite possible there has not even been interbreeding between the Burchell and the other zebras. On the other hand, there is always a chance that any given zebra has been inbred. Horses, on the other hand, with a few possible exceptions, have been interbreeding in all parts of the world; and though sometimes inbred, they are usually supposed to be far less fixed and stable than the zebras. Consequently, on a priori grounds, I expected the hybrid to resemble at least in colour a zebra rather than a horse. I had no very decided opinion as to the shape of the hybrid, but I did not believe what some would-be prophets asserted, that the only result of my experiments would be the production of a monster; that it was little short of sacrilege to cross a good mare with a zebra. Whatever form or colour the hybrid may ultimately assume, I have no hesitation in saying that it would be difficult to imagine any more attractive, more graceful, or more beautiful member of the equine family than the little hybrid Romulus during the earlier months of his existence.
Never ungainly, Romulus when two months old looked as complete and compact as a little horse, and at least twice his age, notwithstanding his being a late foal and the untoward weather that prevailed after his appearance on the scene. This is probably partly due to his beginning to feed on his own account at a very early period. When three days old he was nibbling grass; a few days later he attacked hay, and ere long he insisted on having a share of the oats provided for his dam. Foals frequently content themselves with milk alone during the early weeks.
In shape the hybrid unites the characters of both his parents, and yet without approaching a mule differs from both. When standing on the alert at a little distance, he looks (Fig. 4) as if he had slipped down from a frieze on the Parthenon. The muzzle is very fine, with narrow, almost slit-like nostrils of a distinctly zebra pattern. The forehead is wide as in an Arab, and very slightly convex from side to side. The jaws look narrow, and the head seems to be set on the neck in an uncommon fashion, but the neck is somewhat short. In the neck and its relation to the head, in the position and length of the ears, and in the mane, the hybrid undoubtedly approaches more closely to a zebra than a horse. But beyond the root of the neck I fail to observe any essential difference from a half-bred Arab foal occupying an adjacent box. Both the fore and hind quarters are well formed; the back is short and strong, and the chest is wide and well moulded. In the form of the fore and hind limbs, and in the hoofs, Romulus also resembles in some respects the half-bred Arab, in others his zebra sire; and his tail, with the exception of several bands at the root and the presence of somewhat stiff hairs, agrees with that of an ordinary foal. As in the zebra, there are rudimentary teats, the chestnuts (warts) are absent in the hind limbs, and further there is no tuft of hair at the fetlocks. A little in front of the upright mane there is a separate tuft of hair as in the zebra, and as in many, if not all foals.
The Colour and Striping of the Hybrid. - Darwin, who devoted much time to studying the colour of the horse, came to the conclusion that all the existing races had descended from "a single dun-coloured, more or less striped, primitive stock, to which our horses occasionally revert." [* 'Animals and Plants,' vol. i, p. 65.] In Dzungaria there appear to be still wild horses, i.e. horses whose ancestors have never been domesticated. The wild horses of North America, which are believed to have descended from the Spanish horses of Mexico, were of all colours - black, roan, sorrel, &c.; but the Dzungaria horses are all of one colour. In summer they are dun or sandy coloured; in winter light brown. In all probability the Dzungaria wild horse (E. przewalskii) has not departed greatly in colour from the less remote ancestral horse of the northern hemisphere. The absence of shoulder and leg bands on these wild horses is interesting. We shall never get beyond guessing when the stripes first appeared in the horse group, but it may be assumed that it was long before the days of Hipparion and his other three-toed relatives.
Compared with the even-toed ruminants (oxen, sheep, deer, antelopes, &c.), the odd-toed ungulates suffered heavily during the Tertiary period - only the tapir, the rhinoceros, and the horse families have survived. This being the case, it may be taken for granted that at least in some areas the struggle for existence in the case of the non-ruminating ungulates has been very keen. Bearing this in mind, the existence of stripes becomes intelligible if it be admitted that they counted for something in the battle of life. While admitting that the ancestor of all the horses was "more or less striped," it is well to remember that the only wild horse we are acquainted with (E. przewalskii) has but a faint dorsal band; and that while stripes were presumably useful in races living in wide, fertile, richly populated plains, they may not have been specially useful in the case of the Dzungaria and other races living in remote desert regions. In considering the colour of the hybrid it will be well to remember that when distinct types are crossed the progeny are apt to revert to a more primitive type, to assume some of the ancestral colours.
Taking these and other facts into consideration, I expected the hybrid at birth to be of a dun or bay colour, with distinct dark stripes on the legs and shoulders, and less distinct stripes on the face. Bearing in mind the all but complete absence of stripes in Lord Morton's quagga hybrid, I was not a little surprised when I found Mulatto's foal extremely well provided with stripes all over (Fig. 4), in some respects more richly, if less obtrusively, decorated than any of the zebras. As might have been anticipated, the majority of the stripes were dark brown, while the body colour varied from a light bay colour on the lower part of the legs to a rich orange on the upper part of the face. On the under aspect of the neck and trunk, however, the stripes were indistinct; this was especially true of the ventral mesial band which blended with the dark body colour at each side. When the markings of the hybrid are carefully contrasted with the stripes in the Burchell group of zebras, they will be found to differ very considerably, more especially in the head region and over the loins and hind quarters. In the meantime I shall content myself with shortly describing the disposition of the stripes in the hybrid. The forehead is characterised by having a remarkable series of dark brown narrow bands which alternate with a corresponding series of equally narrow orange-coloured bands. Beginning on a level with the eyes these bands extend upwards, forming a number of graceful arches (Fig. 5) which at once remind one of the forehead of an elaborately tattooed Maori chief. Fourteen pairs of these bands can easily be made out, but the upper ones are interrupted by the tuft of hair which I have named the frontal tuft. Curving downwards from between the eyes towards the nostrils are other bands of a similar width and colour, which serve to decorate the front and sides of the face. These stripes eventually lose themselves in the rich brown hair above the level of the nostrils. The somewhat lozenge-shaped space in the centre of the forehead, i.e. the space between the lateral curved bands just mentioned and the loops above the eyes, is occupied by a U-shaped loop, one end of which bends upwards, while the other, accompanied by a nearly mesial stripe, runs for some distance down the middle of the face. Similar bands (twelve dark and twelve light) extend from the base of the ear obliquely downwards over the jaw. The greater part of the head is thus tattooed with bright- coloured narrow stripes arranged in a quite unique fashion. In having this complex arrangement of narrow dark and light bands over the face, the hybrid differs in a striking manner from his sire, but approaches some of the East African and Somali zebras.

Fig. 5. - Romulus, twenty-seven days old.
Fig 6 - Matopo.
The ears, though zebra-like in shape, are not, as in the zebra, distinctly banded. In a light bay Shetland pony in my possession the tip of the ear is white as in the zebra, but in Romulus the upper third of the ear is of a dark brown colour. This colour extends as a broad band towards the base, where it is interrupted by tan-coloured bands having in the main a transverse direction. The ear is lined with a thick coating of long, fine, bright yellow hair. In a front view the light- coloured ears, with the dark upright mane between them, are almost as conspicuous as the corresponding structures in the zebra.
As is usually the case in zebras, a dark band extends downwards from the withers, to bifurcate at the shoulder, the one limb running forwards to the chest, the other backwards behind the elbow. In front of this, which may be known as the shoulder stripe, are a number of cervical stripes, some of which blend as they run across the neck. At the root of the mane, between the shoulder stripe and the occipital crest, over twenty dark bands alternate with a corresponding number of light ones. The latter are continued some distance into the mane by light brown hairs. The last pair of cervical bands meet, as in the zebra, in the middle line over the sternum, and become continuous with a broad but not very well-defined ventral band, consisting of long dark brown hairs. Running downwards across the trunk, behind and nearly parallel with the shoulder stripe, are nine bands, which take the place of the four or five broad vertical bands in Burchell's zebra. Behind these are a number of oblique bands which curve upwards and backwards, the lower extending across the hind quarters. In the space between the vertical and oblique bands are a number of indistinct narrow stripes, while over the croup and rump there are rows of spots. In having these spots and narrow stripes over the hind quarters and loins the hybrid differs from all the zebras.
It will be interesting to note if there is a like failure to develop distinct broad bands over the hind quarters in hybrids that may afterwards be obtained. The remarkable decoration of the hind quarters is more likely to be due to reversion than the elaborate decoration of the face. It may even have some relation to the dappling so common over the hind quarters in most breeds of horses. There is a distinct dark dorsal stripe, with a yellow line at each side, extending along the spinal ridge to the root of the tail. The dorsal band, narrow in front, expands considerably as it proceeds backwards. The tail, as in the zebra, has several cross-bars at its root. In the region of the shoulder-joint there is again a failure to form distinct stripes, but there are numerous bands across both the fore and hind limbs. Twenty-four bars were counted on the left fore-limb, and thirty-one on the left hind limb. The twelve below the "knee" run obliquely downwards and backwards, and become fainter as the fetlock is reached, while those on the "knee" and the forearm are well marked, and almost form complete circles. In the hind limb the bands above and for a short distance below the hock are extremely well marked, while those on the shank and over the fetlock and pasterns are only represented by faint, short, oblique bars, which fail to reach the inner surface. As in the zebra, there are narrow incomplete "shadow " stripes dividing the broad spaces between the well-marked dark bands of the thigh and upper part of the leg.
Without giving further details it will be evident, with the help of the figures, that the hybrid is elaborately striped, and that it profoundly differs from an ordinary foal. If the figures of Romulus are compared with Figs. 1 and 6 - reproductions of photographs of his sire Matopo - numerous differences in the disposition and in the number of the stripes will be readily noticed. Should the next foal, as was said to be the case in the purely bred second and third foals of Lord Morton's mare, be still more banded, or even provided with a fraction of the bands seen in the photograph, a very good case will have been made out for telegony.
It is still impossible to say what colours the hybrid will eventually assume, but I believe a number of the bands will all but disappear, and that the dark stripes will be separated from each other by bay or dun-coloured spaces. It is quite possible that hybrids bred from light-coloured mares may retain a light body colour and, even when full- grown, have the stripes as distinct as a zebra.
Before leaving the hybrid I may point out that its existence raises a number of interesting questions. In 1808 Frederic Cuvier published in the Annals du Museum d'Histoire Naturelle [* Vol. ii, p. 237.] a note on the mating of an Arab horse with a zebra mare which had previously bred with a male donkey. Unfortunately the mare died some months before the period of gestation was completed. Since then several hybrids have been bred between zebra mares and ponies, but apparently a hybrid between a pony mare and any of the members of the large Burchell group of zebras has not hitherto been obtained. Mules, i.e. hybrids between a jackass and mares, are alike common and valuable; while hinnies, i.e. hybrids between a pony and a she-ass, are rare in England, but comparatively common in Ireland. While hybrids between ponies and zebra mares may not prove specially useful, hybrids between a zebra stallion and ordinary mares (what some would call zebra mules) may have a great future before them. Captain Lugard, who has done splendid pioneer work, more especially in East Africa, recommended some years ago " that an attempt should be made to obtain zebra mules by horse or donkey mares," because he believed such mules "would be found excessively hardy, and impervious to the fly [the dreaded tsetse fly] and to climatic diseases."
Captain Lugard believed "that their export might prove one of the sources of wealth and revenue in the future;" for, as he adds, "everyone knows the paucity of mules, both for mountain batteries and for transport purposes, has long been one of the gravest difficulties in our otherwise almost perfect Indian Army corps." [* ' Our African Empire.' ] Like Mr. Tegetmeier, [* Live Stock Journal, 25th Sept., 1896] I have been assured that zebra mules would be great favourites with the natives in India, and further, it is supposed they might prove invaluable in the West Indies. I have already mated seven different breeds of mares with the zebra, mares varying in size from 11 to 15 hands, and, in fact, enough has been already accomplished to show that zebra hybrids could be produced, suitable, as far as make and size go, for all kinds of work. Further, I had last summer a Burchell's zebra filly that was perfectly docile, in every way as domesticated as a Shetland pony. Hence from a mental as well as from a physical standpoint I fail to see why zebra hybrids should not, if properly handled, prove as serviceable to man as either mules or horses - in some circumstances even more so.
Another question of interest is, will the zebra mules breed inter se, or with horses, zebras, or asses, or any of their hybrids? This is a question which it is impossible to answer until experiments have been made. Because ordinary mules have never been known to breed in England it is argued zebra mules must be infertile. But this does not necessarily follow, for as zebras differ from donkeys, so may zebra hybrids (even in the matter of fertility) differ from mules. Darwin writes, "Many years ago I saw in the Zoological Gardens a curious triple hybrid, from a bay mare by a hybrid from a male ass and a female zebra." ['Animals and Plants,' vol. ii, p. 16] This is a case of an equine hybrid (i.e. a male hybrid obtained by crossing a zebra mare with a jackass) being fertile when mated with a pony mare. Granting the information given Mr. Darwin was correct, this case proves that even in the horse family hybrids may be fertile, and strongly indicates that very careful attempts should be made to induce the new zebra hybrids to breed.
Turning from the zebra hybrid, the next most important fact to chronicle is that, by taking advantage of the knowledge gained as to the habits of zebras, there is now in some respects less difficulty and danger in using Matopo for stud purposes than there is in using many ordinary stallions. Matopo, it may be mentioned, has been trained without applying any severe measures whatever, and without making him less a wild animal than he was on his arrival. By gentle treatment he has become quite tractable. Some days ago I observed the stud groom pulling Matopo's tail with the object of bringing him round to drink. This is one side of the shield; but there is another, for the same groom would be at once attacked, should he interfere when mares are in the vicinity of the zebra. Under such circumstances Matopo is quite regardless of whips or other instruments of a like nature.
In concluding my remarks on the zebras I may mention that, like Frederic Cuvier in 1807, I had no difficulty in mating the five-year-old zebra mare with the Arab horse Benazrek; but unfortunately this mare succumbed during the winter. The two-year-old zebi-a filly which I found so docile and attractive also fell a victim to the dreaded para- site Strongyhis.
In addition to working with zebras I attempted various crosses with donkeys and with an Indian (zebu) bull, but so far I have failed to obtain any results.
In connection with my investigations bearing on polydactylism and on the development of the horse I may mention that I have now in my possession several specimens showing extra digits in the horse and a considerable number of horse embryos. Amongst others I have an embryo for each of the weeks from the third to the eighth inclusive. I have already shown that in a horse embryo fourteen inches in length the second and fourth digits though extremely small, are present [* Journal of Anatomy and Physiology, April, 1894]. I am now in a position to state that at the fifth week the second and fourth digits look nearly as long and as well developed as the third or middle digit, - that, in other words, the horse is for a time tridactylous, the outer digits being relatively nearly as large as in the adult rhinoceros.
In addition to studying the embryos I have been examining the foetal appendages. The most interesting facts made out in connection with these appendages are, first, that the rudiments of the foetal villi make their appearance between the sixth and seventh weeks, and are well formed by the end of the eighth week; and second, that as the yolk placenta dwindles, a complex epiblastic girdle appears externally, while numerous apparently non-vascular villi grow out from the allantois into the coelomic space between the rapidly growing allantois and the yolk-sac.
Postscript. - I find I have omitted to mention that Romulus is Mulatto's first foal, that his eyes are of a bluish- grey colour, somewhat darker than in his sire, Matopo, and that the hocks were for a time marked by narrow longitudinal grooves.
Appendix. - Measurements of Romulus (in inches) when two months old: withers to the ground, 38-and-a-half; shoulder- joint to the ground, 29; elbow to the ground, 26-and-a-half; elbow to the fetlock, 20-and-a-half; cannon bone along outside, 8; circumference at knee, 8-and-a-half; circumference below knee, 5; withers on a line with anterior border of scapula to root of tail, 27-and-a-half; withers to occipital protuberance, 19; chest to buttocks, 35; girth behind fore-legs, 39; length of head from occipital protuberance to end of muzzle, 16-and-a-half; width between eyes where narrowest, 5-and-a-half; where widest, 9-and-a-half; length of ear, 6; stifle to hock, 13; hock to ground, 18; circumference of fore pasterns, 5; circumference of hind pasterns, 5-and-a-half; circumference of shanks, 5-and-a-half; circumference of fetlock-joints, 7-and-a-half; circumference of coronets 7-and-three-quarters.
B. - ZEBRA-HORSE HYBRIDS
THE ZEBRA SIRE OF THE HYBRIDS
During the last two years [* 1896 and 1897] I have bred five hybrids by crossing mares with a Zebra (Equus burchelli var. chapmani). The first hybrid was born on Aug. 12th, 1896; the others were born during the summer of 1897. The dams of the respective hybrids are (1) an Island of Rum pony, (2) a Shetland pony, (3) an Iceland pony, (4) an Irish mare, (5) a cross-bred Clydesdale mare. The sire ("Matopo") of all the hybrids is a handsome 12.2 hands Burchell's Zebra, probably from the Transvaal. As Fig. 7 shows, Matopo is well formed, with powerful legs and, for a Zebra, a fine neck and fairly good shoulders. In his movements he is almost perfect. When trotting, the fore legs move gracefully, without suggesting the hammering action of the hackney; and when galloping he seems to bound along as if without effort, and with but little expenditure of energy.

Fig. 7. - Matopo.
Fig. 8. - Matopo.
If Zebras deserve the ill character they have hitherto borne, Matopo must be an exception to the rule. We are too apt to forget that until Zebras have been under domestication for some generations, it is unfair to judge them by the Horse standard, which after all is not so very high. I have known several perfectly docile captured Zebras, and I have had in my possession a filly (taken when quite young in the Transvaal) which from the first was as docile, tractable, and trustworthy as any pony that was ever foaled. I have refrained from handling Matopo for obvious reasons, yet there is never any difficulty in managing him, unless when he is herding mares, or unusually excited. When in a field with mares he is unapproachable, for, regardless of consequences, he attacks all who venture into his vicinity. Galloping up open-mouthed, uttering his characteristic call, he endeavours to seize intruders by the legs. On one occasion, in a small paddock, he guarded a dozen mares so well, that it took four of us nearly two hours to drive them into their boxes. He is, however, easily upset by unusual noises, and there is nothing that drives him into a state of frenzy so readily as carpet-beating, or that cows him so effectually as a coil of rope. I have often wondered if the rhythmic beating of carpets reminds him of the day when in far-off Africa he lost his freedom; of the time when Boers entangled his limbs to music made by Zulus beating their shields with their assegais.
The more characteristic stripes of Matopo are seen in Figs. 7 and 8. Fig. 8 shows a series of pointed brow arches, some of which end in a frontal tuft nearly two inches in length. Continuous with the frontal stripes are a number of vertical stripes. These stripes extend to the muzzle, the dark skin of which is sparsely covered with short light hairs, except above the nostrils where there are dark brown "nostril patches." There is usually a distinct shoulder-stripe in Zebras, passing downwards from the withers to bifurcate about the level of the shoulder-joint. In fig. 2 (Pl. I.) the shoulder-stripe is double, while in fig. 1 it has blended with a humeral stripe. Between the shoulder-stripe and the occipital crest there are usually twelve cervical stripes, all of which run up into the mane to form, with a corresponding number of white bundles, a series of black and white tufts. Lying between the two upright rows of tufts, and continuous with the dorsal band, is the mane proper, consisting of more or less upright black hairs. The most anterior part of the mane, instead of forming a forelock, extends beyond the level of the ears, and projects forwards at right angles to the long axis of the face. Behind the shoulder-stripe there are, on the left side, five broad, nearly vertical stripes, all but the last reaching the dorsal band above, while all but the first reach the ventral band below. Behind the fifth vertical stripe are a number of broad oblique stripes, with indistinct "shadow" stripes between them. One of these oblique stripes, beginning at the root of the tail, runs forward to pass over the point of the ilium (hip) before bending sharply downwards to reach the ventral band. I have named this the great flank-stripe. Below this flank-stripe a second, having a similar course, may be known as the intermediate flank-stripe. The intermediate stripe is followed by a third, which, starting some distance below the root of the tail, runs obliquely across the quarters to bifurcate over the stifle, the anterior division proceeding towards, but not actually reaching, the ventral band. This may be known as the lower or stifle flank-stripe. These three flank-stripes are equally distinct on the right side, the bifurcation over the stifle being especially evident.
In the space formed by the splitting of the shoulder-stripe are several indistinct arches, and below these arches are the transverse bars of the leg. In some cases this V-shaped space contains portions of seven arches, and the legs may be striped to the hoofs. Below the stifle-stripe there are first oblique and then nearly transverse stripes across the hind leg, with sometimes shadow-stripes between. In Matopo the stripes are indistinct on the lower part of the hind leg, but in many Zebras they become more distinct and relatively broader as the hoof is reached. It will be further observed from the figures (a) that the upper part of the tail is distinctly striped, and that only the lower part of the tail carries long hairs; (b) that though there is a large wart (chestnut) on the fore leg, there is no vestige of a wart on the hind leg, and (c) that there is no tuft of hair at the fetlock.
It may be mentioned that in no two Zebras, or on the two sides (Figs. 7 and 8) of the same Zebra, is the striping alike, that in some cases there are nearly as many shadow as there are ordinary stripes on the neck and body; that even in some Burchell's Zebras there are stripes across the croup and rump which suggest the "gridiron" of the Common Zebra (E. zebra); and that while in summer the dark stripes are nearly black and the light stripes cream-coloured, in winter the dark stripes are occupied by fairly long brown hairs, while the light stripes are made up of equally long white hairs; the light tufts at the side of the mane, however, are white summer and winter. It may be added that Matopo, like the majority of the Burchell group of Zebras, being adapted for a life on the plains, has rounded hoofs and comparatively short ears. He thus differs from the Mountain Zebra (E. zebra), and from his stable companion, a white Arabian Donkey, in which the hoofs are long and narrow, while the ears measure 11-and-a-half in., five inches more than in Matopo.
THE HYBRID "ROMULUS."
The oldest hybrid (Romulus), as already noted, was born on the 12th of August, 1896, the period of gestation being 342 days, - in the mare it is usually from 340 to 350 days. The dam of Romulus was a 13-hands, black Island of Rum pony, lent for the experiments by Lord Arthur Cecil, of Orchardmains, Kent. The well-bred black ponies of the Scottish Western Highlands and Islands, which have long been under observation, form a distinct breed, well adapted in many ways for crossing with Zebras. Their resemblance to Eastern Horses has been accounted for by saying they have descended from sires which escaped from the ships of the Spanish Armada.

Fig. 9. - Romulus (seven days old), and his dam, Mulatto.
Fig. 10. - Romulus: twenty-seven days old.
Fig. 11. - Romulus: one year old.
Romulus, when a few days old, was the most attractive little creature I have ever seen (Fig. 9). He seemed to combine all the grace and beauty of an antelope and a well-bred Arab foal. Instead of, like his sire, looking as if freshly painted for a Lord Mayor's Show, he was faultless in colouring and in the disposition of the stripes, spots, and bands. The body colour was chiefly of a bright golden yellow, while the stripes and spots were of a rich dark brown; but what was especially remarkable was the indescribable sheen of his coat, the dark bands being especially lustrous. A casual glance showed that in the plan of his striping Romulus was utterly unlike his sire, and when a careful examination was made, it became evident that in the number and arrangement of the markings he was not unlike a Somali zebra. As Fig. 10 shows, the brow has been tattooed as if to represent a huge finger print. Instead of the four or five acutely pointed frontal arches of his sire, there are fourteen rounded arches, that remind one of the face of the Somali zebra. Instead of twelve cervical stripes, as in Matopo, there are in Romulus twenty-four cervical stripes, all of which can be traced into the mane. In having so many cervical stripes he seems to be more primitive than even the Somali zebra (in which I have never seen more than fourteen cervical stripes), but closely agrees with one of my zebra mares when the shadow stripes are included. The shoulder stripe bifurcates higher up than in Matopo, and there are seven indistinct arches in the triangular space below the point of bifurcation. Behind the shoulder stripe there are nine (Figs, 9 & 11) fairly distinct vertical stripes instead of five, as in his sire (Fig. 7) . Apparently corresponding to the three flank stripes so often seen in Burchell zebras, there are in the hybrid three stripes in front of the stifle, which first run upwards and then arch backwards to end below the root of the tail (Fig. 9).
Between the last vertical stripe and the great flank stripe are three ill-defined oblique stripes. In the triangular space between the first oblique stripe and the ninth vertical stripe are numerous narrow indistinct lines, some of which proceed towards the ventral band, while others join the oblique stripes. In line with these nearly transverse stripes there were at birth numerous spots arranged in nearly transverse rows over the loins and rump. Now that the hybrid is over a year old (Fig. 11) most of the spots have united to form somewhat zigzag narrow bands, almost identical in their direction with the narrow stripes over the hind quarters of the Somali zebra. On the left side the blending of the spots has advanced further than on the right. Counting from the shoulder stripe to the root of the tail, there are forty-three stripes in the hybrid, - about the same number as in the Somali zebra; in Matopo there are only five transverse stripes behind the shoulder stripe (Fig. 7). It seems to me the blending of the spots over the hind quarters of Romulus goes a long way towards proving that stripes ai-e in many cases first represented by spots or interrupted zigzag wavy lines. Between the stifle or third flank stripe and the point of the hock there are a number of dark bands (between some of which are shadow stripes), while below the hock there are first several distinct transverse bars, and then a number of less distinct oblique lines, right down to the hoof. Similar bars and lines occur on the fore-limb. These leg bars were at birth more distinct than in the zebra sire. Continuous with the mane is a well-defined dorsal band (with a nan-ow yellow band at each side) which extends some distance into the tail. The tail in the hybrid had, at birth, long hairs right up to the root, but, notwithstanding this, there were three distinct bars visible at each side; similar tail bars I have once seen in a horse. Though the ears look long in some of the photographs, they are now relatively very little longer (though rounder at the apex) than in the majority of horses. The nostrils, in their shape, position, &c., are zebra-like, and the eyes and eyebrows may be said to be intermediate; but the eyelashes are long and curved, and quite unlike the short nearly straight eyelashes almost invariably found in zebras and horses. The hoofs of Romulus suggest the zebra more than the horse. They seem to be made of excellent stuff, and to stand a good deal of wear. In his movements the hybrid takes more after his sire than his dam. A few minutes after birth he was rushing about his box, impatient apparently to join the parental troop. What has struck me from the first has been his alertness and the expedition with which he escapes from suspicious or unfamiliar objects. When quite young, if caught napping in the paddock, the facility with which he, as it were, rolled on to his feet and darted off was wonderful. The principal enemy of the zebra seems to be the lion. To escape from the lion, great and sustained speed is not so requisite as a decided and rapid bound when the lion makes his spring, or when he is accidentally met with in the veld. This rapidity of getting out of the way has been strongly inherited by all the hybrids. Zebras, as far as my experience goes, are difficult to handle, not so much because they are vicious or intractable, as because they are afraid. At any moment they may be seized by panic, - when they imagine there is a lion in the path, - and, regardless of consequences, rush, it may be, against a wall or a hedge, or into a ditch, reins and bits counting for little or nothing. In schooling the hybrids this habit will require to be allowed for, and the tendency to bound or rush slowly combated. As it has been completely overcome by careful training in some zebras, there should be comparatively little difficulty in breaking the hybrids. As a matter of fact, Romulus leads anywhere, is perfectly docile, allows his feet to be trimmed and his teeth to be examined, and, when little more than a year old, seemed quite willing to carry a small boy on his back.
I mentioned Mulatto is just under 13 hands, while the zebra sire is nearly 12.3 hands. At birth (August 12th, 1896) Romulus measured 34-and-a-half in. (from the withers to the ground); at two months 38-and-a-half in.; at six months 43 in.; and at twelve months 45i in. The rate of growth has been extremely inconstant, e.g. from the 12th of February to the 12th of April, 1897, he only increased half an inch, [He was weaned on the 14tli of February, and fretted not a little for some time after] and from the 12th of June, 1897, to the 12th of September, 1897, he only increased three-quarters of an inch; [During the greater part of this period he was shedding his old and growing a new coat] but from the 12th of September to the 12th of December, 1897, he increased one and a quarter inches. He now (January, 1898) measures 47-and-a-half in., nearly 12 hands, and the circumference of the fore shank is 6-and-a-half in., the knee being 10 in., and the girth 52-and-a-half in.
The foals of the black Island of Rum ponies are frequently of a mouse-dun colour, with at times an indistinct dorsal band, and a cloudy patch over the shoulder. Usually after the first coat is shed the pure-bred foals are dark brown, and later nearly black, with sometimes indistinct dappling over the flanks and hind quarters. As already mentioned, the body colour of Romulus at birth was chiefly of a yellow tint, the yellow approaching bright orange on the brow, while it approached a straw colour at the muzzle and below the knees and hocks. Under the neck and under the belly the prevailing body colour was dark brown, the ventral band being very indistinct.
The ears were lined with fine bright orange-coloured hairs. When only a month old the hybrid began to shed his foal's coat. The light-coloured hairs began to drop out from the face and neck about the middle of September, and by the end of September he looked considerably darker. The yellow and also the dark brown hairs continued to fall out, except over the back, all through October, and by the middle of November only the orange coloured lining of the ears was left to remind one of the rich coat he wore during the earlier weeks of his life. By the end of November the new coat was established. The bright orange facial bands were replaced by much paler bands, the muzzle was nearly brown in colour, the neck and body intermediate spaces approached a mouse-dun colour, while the lower parts of the legs were of a dark brown tint. From the withers to near the root of the tail the hair was especially long and thick. For a time the hair over the croup and the greater part of the rump was so much longer than the hair around the root of the tail that it looked as if part of the hind quarters had been previously clipped. The new coat consisted of a thick layer of woolly hair from half an inch to nearly two inches in length, and of a less complete coat of stronger hairs, many of which were nearly three inches in length. Near their roots all the body hairs were light in colour, which implies that had the hybrid been clipped there would have been little or no indication of stripes left. In the zebra, on the other hand, the dark pigment extends to the roots of the hair, and hence, however short the hair may be, the binding is quite evident. Recently the skin around the root of Matopo's tail was injured, with the result that the hair, together with some of the epidermis, was shed; but even before the points of the new hair could be detected, the position of the dark bands was perfectly distinct. The skin of the zebra has been described as uniformly black, even under the white bands; but it would be more accurate to say it is of a dark grey colour between the bands and stripes, and nearly black where the bands and stripes occur.
About the middle of March the long hairs began to drop out, and by the end of March they came away in handfuls. As the long hairs were shed from the body, the long hairs were shed from the upper half of the tail, with the result that for a time the tail of the hybrid was little better covered than the tail of his sire. By the end of May all the long hairs - light and dark - had vanished, and early in June the dark and mouse-coloured woolly hairs were coming out. By the 6th of June the dark lustreless winter coat had sufficiently gone around the base of the ears and above the eyes to indicate the colour of the summer coat. All through June and July the process of shedding continued, but by the 12th of August - the hybrid's first birthday - the summer coat was fully established. The dark stripes, which consisted chiefly of strong flattened hairs, looked very prominent. The intermediate bands were of a reddish brown colour over the brow, but elsewhere reminded one of the summer coat of a stag. Taken as a whole, Romulus was very decidedly darker as a yearling than during the early weeks of his existence.
As the long hairs were shed from the body and the root of the tail, numerous hairs dropped from the mane. In an ordinary mule (the foal of a New Forest pony) which I have had for some time, all the long hairs of the mane were shed last summer; but in Romulus, either some of the long hairs were retained, or the new hairs came in before the old ones were lost. At any rate, though the mane was shorter and less bulky and consequently more upright during August, it always consisted of numerous long hairs. At present the mane, which consists of wavy hairs from seven to nine inches in length, tends to fall slightly to one side, - the mane falls slightly to one side in some zebras.
By the middle of September Romulus had again lost not a few of the brighter coloured hairs, and since then he has been getting again gradually darker. Probably because of the extreme mildness of the season the long hairs have already (January) begun to fall out in much the same way as they did last March.
All the experts who have seen Romulus agree in considering him a decided improvement on his sire, and more attractive and shapely than his dam. Having been handled from the first, he is, as a rule, extremely quiet. Occasionally, however, he clearly indicates he has plenty of courage and no lack of speed. At present he is particularly attached to a small thoroughbred mare. When separated from this mare he is sometimes as restless as his sire when upset by some change in his surroundings. Last week a strange horse was galloped in the paddock where Romulus happened to be for the day. The hybrid became excited, and gave an excellent demonstration of his trotting and galloping powers, and of how proudly he can carry himself, and this continued for some time after the intruder left the field. Romulus was recently described by an excellent judge of horses in the Scottish Farmer as "a bonnie colt, with rare quality of bone, . . . and with the dainty step and dignity of the zebra." There is nothing about the hybrids, strange to say, that suggests the ordinary mule or hinny.
THE HYBRID "REMUS."
"Biddy," the dam of Remus, is a three parts bred, 14.1 hands Irish mare. She has been in my possession since 1893, and is now nine years old. Biddy is a bay with black points, but no white hairs anywhere, and Remus is her first foal. She is a very gentle, quiet creature, and has always been in excellent condition, winter and summer alike.
Evidently the zebra, before coming here, had not made the acquaintance of any of his equine relatives. When first introduced to Mulatto, he rushed into a corner with his tail between his legs, and uttered peculiar little sounds strongly suggestive of abject fear. Some of the ponies rushed at him open-mouthed; others deliberately pelted him with their heels. On the other hand, a bay Arab stallion and various mares could not have been more alarmed had he been a tiger, or, when he called "Quacha, quacha," a troop of lions. To give him a chance of discovering what sort of an animal a horse is, I turned him loose one evening with a good-natured but very plucky bay Shetland pony. The pony proceeded to tease the zebra, who very soon began to show fight. He was soon circling round the pony with the object of seizing her legs. For a time the pony was unprepared for this mode of attack, but ere long adopted similar tactics, with the result that the zebra was several times brought to his knees." After a couple of hours the duel came to an end, - the damage being very slight on either side, - and ever afterwards Matopo and "Sheila" were excellent friends.
[* I may mention that when his legs are touched with a rope or stick he almost invariably drops on to his knees, or lies down altogether. This is, I think, the result of his having been periodically thrown before he came here that his hoofs might be looked to.]
But even during the spring of 1896 the zebra was ridiculously timid, and even now a very small demonstration leads him to beat a hasty retreat. Biddy was the first fairly large animal he ventured to approach. One day I tied her up in a court about forty feet square, a cloth having been previously bound over her eyes. The zebra in course of time ventured within a few yards; later he laid his head across her quarters, and then, for quite a long time, across her withers. He next licked her lips, and ended by gently nibbling at her ears. Evidently at length satisfied a big horse was after all not so terrible an object, he retired to his box and finished his corn. Having once learned the peculiarities of a mare, he never forgets them. Some of the mares he dislikes, while he is very fond of others, getting quite excited when they pass his own particular quarters. Donkeys, however, he completely refuses to take the smallest notice of.
Remus - born on the 18th May, 1897 - was, at birth, relatively smaller and far less active than Romulus; the period of gestation was three hundred and forty-six days. When a day old he measured 35-and-a-half in., his girth being 28 in. On the 18th of June he had increased to 38-and-three-quarter in., the girth being 36 in. When six months old he measured 44-and-seven-eighth in., the girth being 47-and-a-half in., the circumference at the knee 9-and-three-quarter in., and below the knee 5-and-three-quarter in. Romulus at six months old was 42 in.
From the first Remus has been extremely friendly, and yet in some respects he is more zebra-like than Romulus. For some days he was little more than a machine, - an automaton capable of following a moving object and of sucking. All the special sense-organs were apparently at work, but the brain seemed incapable of making much use of the information collected. If I moved away he followed me, and sucked at my fingers or anything else offered him. He heard his dam when she called, but he was unable to discover whence the sound came, and when he saw her at a few yards distance he failed to recognise her. He seemed to like aloes and water quite as much as sugar and milk, and did not mind either strong smelling-salts or freshly-made mustard. Though he kicked aimlessly when pinched, he paid no heed to the application of either warm or very cold substances to the skin. When a dog was first introduced to Romulus, his excitement was intense. He rushed about at a furious rate, striking as opportunity offered with his fore-feet, holding his head high and stepping high, as if moving through long grass, where other enemies might lie concealed. Remus, on the other hand, when two days old, allowed a yellow collie to lie down within six inches of his muzzle, and only got up as a Dalmatian approached, when a warning note was uttered by hiswvatchful parent. When the four 1897 hybrids and two pure-bred foals were eventually weaned, Remus seemed to mind very little. While one of the hybrids and a half Arab foal were biting and kicking and rushing about as if demented, Remus simply stood looking over the fence. But by-and-by, when the others settled down, he set to walking backwards and forwards behind the wall of his court, exactly like his zebra sire, and though he still keeps this up as if he were a caged lion, none of the others have followed his example. When Romulus was weaned, he for some days rushed about, as much like a zebra when highly excited, as his sire when upset by the beating of carpets. Recently it was necessary to give the hybrids milk containing thymol. The pure-bred foals offered but little resistance, but all the hybrids fought till they were exhausted, and nothing would persuade Remus to swallow the first dose.
As might have been expected, Biddy's foal is much lighter in colour than Mulatto's. With the exception of the muzzle and the lower part of the legs, the body colour is a rich light bay; the muzzle and legs were, at birth, more of a mealy colour, but are now of a bay colour. The bands are much lighter, and consequently less distinct than in Romulus. As a rule they are of a dark reddish hue, being especially evident on the brow, the forearms, and above and below the hocks. The plan of the striping is the same as in Romulus; but even at birth several of the rows of spots across the croup had already united to form narrow bands. The face, measured from the occipital crest to a line connecting the upper margins of the nostrils, was slightly longer than in Romulus; but the ears were the same length - six inches.
Sometimes when a horse utters a warning call all the members of the herd hurriedly collect together and rush about in an excited manner. It seems to be of the utmost importance for wild Equidae to at once make out the direction of any given sound. Probably the longer the ears the quicker this is accomplished. If the length of the ears, as is most probable, counts for much, one can understand why they almost reach their full size at birth. Foals are given to straying in all directions, and unless they hear and at once recognise the call of their respective dams, and the direction from which the sound comes, their chances of surviving in a wild state would be greatly reduced. At birth, the ears of Romulus were longer than in his dam, and only slightly shorter than in his sire. In the case of Remus they were the same length as in his dam, viz. six inches along the upper aspect.
The eyes in Biddy's foal are hazel-coloured and gazelle- like in their mildness, and the upper eyelashes are particularly long and curved. The mane, though at first made up of soft hairs, which bent over to the right side, soon assumed an upright position, and when Remus reached the eighth month, it consisted of nearly erect but not very stiff hairs. It looks as if the mane will always be as upright and as short as in his sire. The tail contains fewer hairs than any of the other hybrids, and has three bars across the root. On the other hand, there are chestnuts on the hind legs as well as on the fore; ordinary mules are said not to have chestnuts on the hind limbs. The front chestnuts are large, level with the skin, and zebra-like; the hind chestnuts are raised above the level of the skin, and, though narrow and only half an inch in length, are horse-like. That the zebras and asses have no chestnuts on the hind legs may perhaps be due to the absence of chestnuts in their remote ancestors; their absence points, I think, to asses and zebras having sprung from a different ancestor (perhaps Hipparion) than the horses, which may have descended straight from Protohippus, If Remus survives, he may reach a height of nearly fourteen hands, and be the most handsome and fleetest of all the present crop of hybrids.
As in the case of zebra foals, the hair over the back and hind quarters of Remus soon increased in length, and formed a thick woolly covering. The hair of the first coat usually falls off soonest from the face and neck, then from the legs, especially at the knees and above and below the hocks. Some of the hair was shed from the face by the end of the first month, but there was still some left on the muzzle and brow at the end of the third month, and the legs retained some of the foal's coat at the end of the fourth month. The second coat, which was completed by the end of' the fifth month (i.e. about the middle of October), consisted of a thick inner coat of bay and brown fine wavy hairs, averaging an inch and half in length, and of an outer but much less abundant coat of stronger hairs, many of which were two and a half inches in length. Neither the long nor short hairs nor the hairs of the mane have yet (January) begun to fall out.
THE HYBRID "BRENDA."
The dam ("Lady Douglas") of Brenda is a cross-bred Clydesdale mare, built on the lines of the "Douglas" breed once common in the Hamilton district. Like Biddy, she is a bay with black points, but, unlike the Irish mare, she has a large "blaze" on the face, a heavy mane and tail, and a liberal amount of hair at the fetlock-joints. Lady Douglas is fifteen hands high, the circumference at the knee is thirteen and a half inches, and below the knee nine inches. The face is longer than in Biddy by nearly an inch, and the ears by three quarters of an inch. I expected Brenda (the Clydesdale's first foal) to closely resemble Remus in colour and markings; but in breeding, more especially in cross-breeding, the unexpected often happens. We are too apt to forget that, even when the sire belongs to a different and very distinct species, the progeny may take after the cross-bred dam. It was evident soon after Brenda (Fig. 12) was foaled that she differed not a little both from Romulus and Remus. In the first place her ears looked extremely long; they were at birth six and a half inches, only a quarter of an inch shorter than the ears of her dam, and quite as long as those of her sire; at six months they were seven and a half inches. On the other hand, the head is relatively short - shorter than the head of a twelve-hands Iceland pony's hybrid. The height at the withers was forty-three inches, one inch more than in Remus, and four inches more than in the Iceland hybrid.

Fig. 12. - Brenda: two months old.
At birth Brenda, apart from her ears, looked not unlike an ordinary bay foal, but soon faint stripes began to show themselves, and in a day or two the stripes, though indistinct, were seen to closely agree in their arrangement with those of the other hybrids. Now that the "Clydesdale" hybrid is nearly seven months old, she at a little distance might easily be mistaken for an ordinary foal. Compared with Remus the head is shorter and finer, while the joints are larger and the shanks thicker. At six months the circumference at the knee was ten and a quarter inches, and below the knee six and one eighth inches - almost exactly the same as in Romulus when seventeen months old. The mane, at first nearly upright, short and zebra-like, is now made up of hairs from eight to ten inches in length (nearly as long as in an ordinary foal of the same age). Except near the withers and between the ears the mane arches freely to the right side, some of the hairs almost touching the neck. The hair between the ears already projects forwards to form a forelock. In Remus, as already mentioned, the mane is still upright, and shorter than in his sire. The tail in Brenda has also from the first been heavier than in any other of the hybrids, and fewer hairs have been shed from its base; farther, almost from the first there have been a few hairs at the fetlock-joints. The hairs around the small ergots are now over two inches in length.
The chestnuts on the fore-legs in the zebra are large and smooth, and on a level with the skin; in Romulus and Remus they are also large, and hardly if at all above the level of the skin, but they occasionally give off thin scales. In Brenda the front chestnuts, though relatively nearly as large as in a zebra, project nearly as far above the level of the skin as in a pure Clydesdale foal. The left hind leg carries a small prominent chestnut about a quarter of an inch in diameter, but there is no rudiment of a chestnut on the right hind leg. The hoofs are the hoofs of a zebra, and considerably smaller than would be the hoofs of a Clydesdale foal of the same age. They are wide behind and rounded in front, but the bars are relatively short, i.e. they do not extend as far back as the frog. I may add, the nostrils are in their shape a little less zebra-like than in the other hybrids; that the muzzle suggests the dam more than the sire, the lower lip being, as in the dam, somewhat long; and that the rounded ears are tipped with white, as is occasionally the case in dun ponies as well as in zebras. As might have been expected, the trunk and hind quarters are more massive than in Remus, while the shoulders are less upright, and perhaps as a consequence of this the action at all times is less zebra-like than in any of the other hybrids. As Fig. 12 indicates, there is a "swirl" nearly three inches in length extending down the centre of the face between the eyes. The same figure also indicates fairly well the extent of the marking at the end of the second month. The brow arches (hardly visible in the figure) are nearly as pointed as the frontal arches in a Norwegian pony in my possession, and as in the Amsterdam quagga. This is very remarkable, as in all the other hybrids the brow stripes form rounded arches. The cervical, and in fact all the other stripes as far as they go, agree with the corresponding- stripes of Romulus. In the region of the shoulder the markings are very faint, and over the hind quarters only a few indistinct spots and portions of bands can be detected. The lower part of the legs is only faintly striped, and even the bars across the forearm and the hock are more obscure than usual. But although none of the stripes are very pronounced, there are, strange to say, faint lines between several of the cervical and vertical body stripes. These lines suggest "shadow" stripes, and seem to correspond to some of the numerous indistinct vertical stripes seen in zebra-ass hybrids. In having faint intermediate vertical stripes, this, on the whole, horse-like hybrid may be said to be, in at least one respect, more primitive than either of the other hybrids already described. If this hybrid continues to thrive, she ought to grow into a powerful, active, shapely cob, about fourteen hands in height, hardier and with more staying power than an ordinary mule.
THE HYBRID "NORETTE."
The most attractive of last summer's crop of hybrids has for its dam a good-looking eleven hands Shetland pony ("Nora"). This pony, which will be sis years old in the spring, had a foal in 1895 to a small black Shetland pony ("Wallace"). Nora is in many ways a small edition of Mulatto, and her foal Norette may be said to be a small edition of Romulus. When a few days old Norette, in her colouring, movements, and make, was more fascinating than Romulus at a similar age; and now that she has increased from thirty inches, (her height when foaled on June 8th) to nearly forty-one inches she looks as if she belonged to some bygone age. Norette has been from the first more intelligent than any of her contemporaries, and always very much on the alert without being at all nervous or frightened. She followed her dam through a crowd of some thousands of people on Jubilee Day without any hesitation or evincing any signs of fear, and she now leads quietly and allows herself to be measured without offering any resistance. At birth Norette generally resembled Romulus, both in colouring, markings, and shape, but her head was relatively smaller, and the ears relatively shorter. There was, however, a very important and interesting difference between Norette and the other hybrids. As already pointed out, the croup and rump of Romulus were at the outset marked by numerous rows of spots having on the whole a transverse direction. When his new coat was completed, in August last, I noticed that many of the spots had united to form somewhat zigzag bands which in their direction agreed closely with the stripes on the hind quarters of the Somali zebra. In Norette, instead of spots over the hind quarters, there were from the first numerous narrow and hardly at all wavy stripes, which line for line almost agreed with the markings in the Somali zebra. But further, many of these all but transverse stripes reached, or all but reached, a stripe running obliquely across the hind quarters in almost the same position as the oblique stripe in the Somali zebra, which I have elsewhere referred to as the upper femoral stripe. The remarkable difference between the markings over the hind quarters of Norette and her sire Matopo, and the equally remarkable resemblance between these markings in Norette and the Somali zebra, seem to me to throw a flood of light on the relationships of the stripes in the various species and varieties of zebras, and at the same time strongly to support the view already advanced, that the difference between the stripes of the sire and his various hybrid offspring is in all probability due to atavism or reversion. If this is the correct explanation, it follows as a matter of course that, at least in the markings, the Somali is the most primitive of all the known recent zebras.
That the hybrids have reverted in at least their markings towards a somewhat remote ancestor of the zebras is also indicated by the presence of faint "shadow" stripes on the neck. From Matopo having twelve cervical stripes and some zebras having in addition nine or ten " shadow " stripes, and from Romulus having twice as many stripes as Matopo, it may be inferred the typical number of cervical stripes in zebras is twenty-four or thereabout. But in Norette, in addition to the twenty-four cervical stripes there were at least five faint "shadow" stripes. In zebra-ass hybrids there are usually many indistinct stripes on the neck and body, and numerous spots over the hind quarters. I consider zebra-ass hybrids more primitive in their markings than zebra-horse hybrids. In having numerous cervical stripes Norette approaches zebra-ass hybrids, and the only explanation of this that occurs to me is that in Norette we have, in the striping of the neck, a further reversion than in any of the other hybrid offspring of Matopo,
During the first three months the mane of Norette was quite upright, though thicker than in the other hybrids. During the last four months the mane has been increasing in length, aucl it is now no longer upright; the posterior half hangs over to the right side, the part between and in front of the ears forms a thick forelock, while the intermediate portion hangs to the left side.
Norette with her short head, peculiarly tattooed face, and the heavy mane hanging partly to one side and partly to the other, looks very quaint, and seems to differ quite as much from her sire as she does from her dam the black Shetland pony. At seven months the coat was very heavy, the long hairs of the body measuring over three inches, while the hairs over the brow were nearly two inches in length. If Norette develops after the fashion of Romulus, she will - a year hence - be a compact small striped pony from 11 to 11.2 hands in height. As is the case with Romulus, there is nothing about Norette that suggests either an ordinary mule or a hinny. She has excellent, well-formed feet, only a few short hairs at the fetlock, and not a rudiment of warts on the hind legs.
THE HYBRID "HECKLA."
Heckla's dam is a twelve hands skewbald Iceland pony. There is so much white in this pony (Tundra), and the yellow is so pale, that I thought her hybrid foal would be nearly as light as a pure-bred zebra. As it happens, Heckla is the darkest of all the hybrids, and the stripes were at birth as obscure as in the "Clydesdale" hybrid Brenda. As she lay by her dam shortly after birth, she looked like an overgrown hare with an unusually long-head and relatively long ears. From the first her coat has consisted of long coarse hairs, and the warts on the front legs are prominent, as in her dam. Measuring 32-and-a-half in. at birth, she was 43 in. at six months, and is now (January 12th) 43-and-a-half in.; the circumference of the knee being 9-and-a-half in., and the fore-shank 5-and-a-half in. Though Heckla has always carried a heavy coat, and is dark in colour with light tips to her ears, she generally agrees with Romulus in her build and markings; but her action is freer, and more like that of a hackney than a zebra. She promises to be quite as large and as active as Romulus, and more able than Romulus to withstand cold and to flourish under adverse circumstances.
The length of the head and the shortness of the neck suggest that the Iceland ponies do not belong to the same race as the black Oriental-looking West Highland ponies. They may be direct descendants of the horses hunted by the men of the Reindeer Period. Their ancestors may have gradually worked their way northwards with the Tundra fauna, which then, as now, lived near the edge of the ice. If Heckla owes her dark colour to reversion, it may be inferred her ancestors were of a mouse-dun colour.
It is too soon to offer any opinion as to whether Romulus or any of the zebra-mare hybrids will prove fertile or specially useful either at home or abroad, and it is equally impossible to say whether they will withstand the African tsetse fly, or have better constitutions than either ordinary mules or asses; but this much may be said, they all seem very hardy. Romulus has been in perfect health from the first, as indeed has been his zebra sire, while nearly all my mares and horses have had colds and other ailments. Quite recently the four hybrid foals and three ordinary foals have been suffering from the presence of various species of Strongylus. One of the pure-bred foals (Mulatto's second foal to an Arab horse) died from the effects of the parasite on the 1st of January, and a thoroughbred foal has been reduced almost to a skeleton; but the four young hybrids, though no longer so bright or in so good condition, are evidently rapidly recovering, and will, I trust, be soon all right again. I may mention that the editor of the Scottish Farmer believes Romulus "will be invaluable for driving or riding on account of his hardiness," and he has stated that all the hybrids "have feet and legs like whalebone, with the kind of pasterns that Clydesdale men fancy." It is well known that Captain Lugard and Major von Wissmann have advocated that steps be taken to breed zebra hybrids. [* Scottish Farmer, November 27th, 1897.]
I have already referred to the views of Captain Lugard. He writes: - "Some years ago I advocated experiments on taming the zebra, and I especially suggested that an attempt should be made to obtain zebra mules by horse or donkey mares. Such mules I believe would be found excessively hardy and impervious to the 'fly' and to climatic diseases I would even go further and say that their export might prove one of the sources of wealth and revenue in the future; for, as everyone knows, the paucity of mules both for mountain batteries and for transport purposes has long been one of the gravest difficulties in our otherwise almost perfect Indian Army Corps." Since this was written much information has been gained as to the dreaded tsetse fly, but apparently there is extremely little chance of horses being made immune, being so treated by inoculation or otherwise that they will be able to survive if once infected by the peculiar minute organism so intimately associated with the all too fatal disease.
Further, owing to the destruction of cattle by the rinderpest, the transport difficulties have been increased in Africa, while the frontier wars have enormously increased the demand for mules in India. On the other hand, it has been proved that it is a comparatively simple matter to cross various breeds of mares with a Burchell zebra, and if experts are to be trusted the hybrids (zebra-mules as some call them) promise to be as useful and hardy as they are shapely and attractive. The preliminary difficulties having been overcome, it remains for those in authority to take such steps as may be necessary to ascertain of what special use, if any, zebra hybrids may be in the various parts of the Empire, but more especially in Africa and India.
As I am anxious to obtain as much information as possible bearing on equine hybrids - on crosses between zebras, horses, and asses - and as to the fertility of the various kinds of hybrids (mules, hinnies, &c.), I shall be most grateful for accounts of any experiments hitherto made, more especially with Burchell and other zebras. I have not yet heard of ordinary mares having been crossed with Burchell's zebra in South Africa; but doubtless some of the readers of The Zoologist may be able to give me information on this subject.
PART II. TELEGONY AND REVERSION.
TELEGONY, WITH OBSERVATIONS ON THE STRIPING OF ZEBRAS AND HORSES, AND ON REVERSION IN THE EQUIDAE.
INTRODUCTORY.
Of the still surviving ancient and widespread beliefs, one of the most suggestive and interesting to students of nature is the belief that parents hand on to their offspring some of the peculiarities acquired during their lifetime; in other words, that acquired characters or modifications - characters not due to heredity - may be transmitted. This is a tradition likely long to withstand the scotchings of science, even should it, like so many other traditions, be eventually found to rest on foundations of sand. The transmission dogma may be said to consist of three main stems or trunks, which are often associated with, and may be named after, the patriarch Jacob, Lamarck, and Lord Morton; the heads borne by these trunks are, however, in the meantime best left unnumbered and unnamed.
The Jacobean Trunk. - I have frequently heard of Aberdeenshire farmers who acted as if they believed that were peeled rods of hazel, poplar, or chestnut placed before their cattle they would bring forth young ring-straked, speckled and spotted." Amongst shepherds the belief is prevalent that if a single black ewe is allowed in the autumn to run with a flock of white ones, however perfect their pedigree, a considerable percentage of the next crop of lambs will be black. Whether the methods at times practised by breeders in various parts of Scotland to secure a desired end are the result of observation and experiment, or are due to a Boer-like faith in the teaching of the Old Testament, I am unable to say. But assuredly breeders of Aberdeen-Angus cattle have not been singular in believing in the power of mental impressions on the offspring. The tradition influenced the ancient Greeks; it still holds sway in modern Rome.
The Lamarckian Trunk. - In 1801 and again in 1809 Lamarck ventured to suggest that the various species of animals had not (as was then all but universally believed) been separately and specially created. Like Erasmus Darwin, he believed that from a few simple forms all the varieties and species of animals had been gradually evolved. He accounted for the progressive increase in organisation in the vegetable and animal kingdoms by saying that as new organs were needed they appeared - partly through transcendental influences, - and that the modifications acquired by one generation were handed on to the next. In other words, Lamarck believed that the evolution of plants and animals had been mainly accomplished by the transmission of acquired characters, by the inheritance, generation after generation, of functionally produced modifications, together with changes resulting from the influence of the surroundings or environment. To give the familiar examples, Lamarck believed that the necks of the ancestors of the giraffe had been lengthened by generation after generation stretching them to reach the higher foliage; that webbed feet had been acquired in, e.g., aquatic birds and mammals, by the toes being outstretched when swimming; and, to take another example, Lamarck believed that certain four-footed reptiles, by wriggling amongst shrubs and grasses, by elongating their bodies and losing their worse than useless limbs, had been gradually transformed into snakes, while others by acting in a different fashion had developed into birds. However incredible it may to some appear, Lamarck's theory in a modified shape still forms part of the creed of not a few biologists in Europe and of a still larger number in America. Some of the Neo-Lamarckians may, like Lamarck, believe that improvements partly result from the operation of some transcendental principle working in predetermined lines - believe in a kind of teleology.
[* It will, I think, be allowed that most people who come into touch with nature, whether they admit it or no, are more or less teleologists; besides admitting that there are unfathomable mysteries, they, unconsciously it may be, believe not only that there is order in the universe, but that a kind of intelligence pervades nature.]
The Mortonian Trunk. - For all we know, breeders since the days of the patriarchs may have believed not only in the influence on the offspring of mental impressions, but also in the influence of a previous sire; that the offspring sometimes resemble not so much "the father, but an early mate of the mother," and that the female comes to resemble more and more the male to which she continues to bear offspring. We certainly know that what used to be spoken of as the "infection of the germ," but which, following Weismann, we now-a-days call "telegony," [* from the Greek "at a distance;" and " offspring."] was considered possible by physiologists at the end of the seventeenth century; we know the infection tradition has long influenced Arab breeders, and that believers in this hypothesis may now be found in every part of the world, more especially wherever an overlapping of distinct races occurs, as, e.g., in the Southern States of America and in certain Turkish provinces. Further, until quite recently many biologists considered that what is commonly and conveniently known as Lord Morton's experiment had proved "infection of the germ" to at least occasionally take place. Of these three main trunks I propose dealing with the last, i.e. with telegony, and with some of the results obtained by crossing zebras and horses.
LORD MORTON'S EXPERIMENT, AND THE VIEWS OF DARWIN, WEISMANN, AND OTHERS.
It is admitted on all hands that the time has more than come when the belief in the transmission of acquired characters should be tested by carefully conducted experiments. Already a number of isolated experiments have been made without any decisive results, but (probably on account of the difficulties to be overcome) no one has hitherto repeated Lord Morton's experiment. Weismann, who maintains that natural selection has been the only influence at work in the evolution of all except the simple unicellular organisms, has shown that the evidence in favour of the transmission of acquired characters is in many cases worthless, and in no case convincing. [See also Platt Ball, 'The Effects of Use and Disuse,' 1890] In discussing the "infection" theory in 1893, Weismannf pointed out that new experiments after the manner of Lord Morton's were needed, and that they could "be best made in zoological gardens." ['The Germ-plasm' (Contemporary Science Series). Perhaps the time may come when systematic experiments of this kind will be one of the characteristic features of the Regent's Park and other zoological gardens, or when an experimental station will be founded in memory of Darwin.]
Having failed to induce zoological or other societies to take part in repeating Lord Morton's experiment - work which it was generally admitted should be undertaken without further delay, - I decided in 1894 to start on my own account a number of investigations bearing more or less directly on telegony. I knew that numerous experiments on this subject had yielded negative results, and that some biologists held that such inquiries were all but useless. On the other hand, I believed that by working with zebras and horses I should obtain more decided results, one way or the other, than with any other mammals, and that, apart from any possible scientific gain, it was worth while for various reasons attempting to produce hybrids be- tween zebras and certain breeds of horses. [There seems little chance of rendering horses immune to the bite of the tsetse fly, but zebra hybrids may withstand it; those I have already bred seem uncommonly hardy. Further, hybrids between zebras and English or other mares may be found invaluable in various parts of the Empire. I have one that looks as if it would by-and-by be well adapted for a mountain battery.]
I now propose stating shortly the results of the experiments, as far as they directly or indirectly bear on telegony and on reversion or atavism. But before doing this it will be advisable to discuss Lord Morton's experiment, and refer to the views held by Darwin, Spencer, Weismann, Romanes, and others on the "infection" theory.
Lord Morton's object apparently was not to ascertain whether there was such a thing as " infection of the germ," but simply to see whether it was possible to domesticate the quagga, now, alas! extinct. In the letter addressed to the President (Dr. Wollaston) of the Royal Society on August 12th, 1820, Lord Morton says: "I obtained a male, but being disappointed of a female [quagga] I tried to breed from the male quagga and a young chestnut mare of seven-eighths Arabian blood, and which had never been bred from. The result was the production of a female hybrid, now five years old, and bearing both in her form and in her colour very decided indications of her mixed origin. I subsequently parted with the seven-eighths Arabian mare to Sir Gore Ouseley, who has bred from her by a very fine black Arabian horse, ... a two-year-old filly and a year-old colt, which in their colour and in the hair of their manes have a striking resemblance to the quagga." [* 'Phil. Trans.,' 1821, p. 21. For complete letter see Appendix.] It is further mentioned in the now famous letter that the colt and filly were in some respects more striped than either the quagga or the quagga hybrid, and it is added that while the mane of the filly was stiff, short, and erect, as in the quagga, that of the colt arched upwards, as in the hybrid.
Notwithstanding these quagga or hybrid-like characters, few biologists at the present moment admit that Lord Morton's chestnut mare was "infected" by the quagga. Darwin, however, in 1875 seems to have believed the chestnut mare had been "infected." He says, "These colts [the colt and filly bred by Sir Gore Ouseley] were partially dun-coloured, and were striped on the legs more plainly than the real hybrid or even than the quagga. One of the two colts had its neck and some other parts of its body plainly marked with stripes. Stripes on the body, not to mention those on the legs, are extremely rare - I speak after having long attended to the subject - with horses of all kinds in Europe, and are almost unknown in the case of Arabians. But what makes the case still more striking is that in these colts the hair of the mane resembled that of the quagga, "being short, stiff, and up- right. Hence there can be no doubt that the quagga affected the character of the offspring subsequently begot by the black Arabian horse." [* 'Animals and Plants under Domestication,' vol. i, p. 435, 1875.]
In the same chapter Darwin, referring to a horse bred by Lord Mostyn from a mare that had previously borne a foal to a quagga, says, "This horse is dun with a dark stripe down the back, faint stripes on the forehead between the eyes, plain stripes on the inner side of the fore-legs, and rather more faint ones on the hind legs with no shoulder-stripe. The mane grows much lower on the forehead than in the horse, but not so low as in the quagga or zebra. The hoofs are proportionally longer than in the horse, - so much so that the farrier who first shod this animal, and knew nothing of its origin, said, 'Had I not seen I was shoeing a horse I should have thought I was shoeing a donkey.' " It is especially worthy of note that Darwin favoured the view that Lord Morton's mare had been infected, notwithstanding the fact that he had bred and described a colt which for some weeks after birth was marked over the body, legs, and face by numerous narrow stripes. [Loc. cit., vol. i, p. 60.] Many cases of supposed infection in birds, and still more in domestic mammals, have been put on record, but none of them are in my opinion so striking as the two just mentioned.
One of the most industrious collectors of supposed cases of telegony was Dr. Harvey of Aberdeen, who contributed papers on this subject to the Monthly Journal of Medical Science for the years 1849, 1850, and 1854, and published in 1851 a pamphlet on 'A Remarkable Effect of Cross-breeding.' Harvey eventually persuaded himself that there was such a thing as "infection." He believed he had finally raised the tradition to the rank of a fact, and accordingly discussed at some length the possible methods by which the "infection" was brought about.
Of quite recent writers, Herbert Spencer has felt most satisfied as to the fact of telegony. In a series of papers in the Contemporary Review [Vol. lxiv.], in which telegony has practically been taken for granted, Spencer has especially concerned himself with finding an explanation of the phenomenon. Spencer thinks that the evidence he submits by "coming from those who have no theory to support, dissipates all doubt," and he endeavours, as will be further explained below, to account for telegony by supposing that germ-plasm from the embryo reaches and "infects" the maturing ova through the tissues of the dam.
The late Mr. Romanes (whose telegony experiments were abruptly cut short by his untimely and widely lamented death), in a reply to Mr. Herbert Spencer says, "On the present occasion space does not admit of giving special instances, so I must ask it to be taken for granted that my evidence is enough to prove the fact of a previous sire asserting his influence on a subsequent progeny." [Contemporary Review, April, 1893.] Elsewhere, on giving the upshot of an extensive correspondence on the subject under consideration, Romanes states that "the principal result is to show that the phenomenon is of much less frequent occurrence than is generally supposed. Indeed, it is so rare that I doubt whether it takes place in more than one or two per cent, of cases." ['An Examination of Weismannism,' 1893, App. 2, p. 199.]
Before stating the view Weismann holds as to "infection" I perhaps ought to indicate what English breeders think about the subject. This will be best accomplished by the following quotations from two letters which appeared in The Sportsman early in 1896. The writer of the letters (the "Special Commissioner " to The Sportsman) is, I understand, one of the most influential men in racing circles in England. In a letter dated January 3rd, 1896, the "Special Commissioner" says, " There would seem to be gentlemen who are as yet unaware that previous matings do undoubtedly in many cases exercise an influence on all the subsequent stock of a mare or other female animal. Now I should never have expected to find anyone disputing this undoubted fact, and the only question appeared to be whether the old theory, which ascribed it to the imagination of the mother or some equally unsatisfactory cause, should not be replaced by the infinitely more sensible doctrine of saturation, which is thus defined by Mr. Bruce Lowe: - 'Briefly put, it means that with each mating and bearing the dam absorbs some of the nature or actual circulation of the yet unborn foal, until she eventually becomes saturated with the sire's nature or blood as the case may be.' Bruce Lowe, ['Breeding on the Figure System.'] ... I confess that this theory has always appeared to me as a most reasonable one. As to the fact of sire influence which it professes to explain, I may inform correspondents who are in any doubt about it that it would not be difficult to furnish them with hundreds or even thousands of instances" [The Sportsman, January 3rd, 1896.] In a subsequent letter (January 22nd) the "Special Commissioner" says, "I should as soon have expected anyone to boldly contend that the earth is not round ... as to deny the existence of this influence," i.e. of the influence of the previous sire. Another writer in the same paper says, "It is perfectly surprising that anyone should doubt that saturation is an important part of the theory of breeding. There would be little difficulty in showing that it is not only a fact, but a proven and most important factor." This writer tells us an experienced veterinary surgeon, who also raced, often said to him, "It is of the greatest importance to mate your mare the first time with a good horse; she will always throw a bit towards him."[Ibid., January 17th, 1896.]
Similar advice would be given by the majority of veterinary surgeons if they hold with the view that emanated some years ago from the Principal of the Bombay Veterinary College. In an interesting paper on mules the Principal says: - "An extraordinary influence is exerted on the system of a mare by becoming the mother of a mule, in so far as her progeny in the future is liable to a taint indicated by asinine characters cropping up. Thus no brood mare of high value should be made to throw a mule - a fact well established by the experiment made by Lord Morton of crossing a mare of pure blood with a quagga." [Mr. J. H. Steel, A.V.D., ' Journ. of the Bombay Nat. Hist. Soc.,' vol. V, 1890.]
It is, perhaps, not to be wondered at that in England, where there has been hitherto in many quarters a dread of scientific methods akin to that of the supernatural, breeders have failed to distinguish between facts and beliefs. In this case inquiry may justify the wide acceptance of the belief in question, but until the verification has been effected it is not wise to allow the tradition to overmuch influence practice, or to argue, as some believers in telegony have done, that because the dam is infected the sire must be infected also; that a Chillingham bull, e.g., which has been used for breeding with shorthorn cows will, if returned to his own herd, infect it with shorthorn peculiarities and idiosyncrasies. While many English breeders have been, it may be, over-credulous, not a few German breeders have long looked with suspicion on the infection theory. Professor Kuhn, late head of the Prussian Agricultural Station at Halle, Settegast, Nathusius, and others familiar with scientific methods have, notwithstanding an extensive experience in breeding and crossing, never known a case of telegony. Hence, while some have doubted its ever occurring, others are convinced there is no such thing as telegony, that the female is neither "infected" by the first male nor by subsequent mates to which she bears offspring.
Weismann, who, as is well known, is not a believer in the infection theory, wrote in 1893 as follows: - "I must say that to this day, and in spite of the additional cases brought forward by Spencer and Romanes, I do not consider telegony has been proved. I do not dispute the possibility of telegony; I grant that the wide general acceptance of the belief in the past has so impressed me that I have always said that possibly it might be justifiable and founded on fact. I should accept a case like that of Lord Morton's mare as satisfactory evidence if it were quite certainly beyond doubt. But that is by no means the case, as Settegast has abundantly proved. . . . The attempt must be made to determine the truth by new experiments. . . . According to scientific principles, only the confirmation of the tradition by methodical investigation, in this case by experiment, could raise telegony to the rank of a fact." [Contemporary Review, vol. Ixiv.] Not a few biologists and breeders agree with Weismann in questioning the fact of telegony, in considering "infection" as extremely improbable if not absolutely impossible. Why, it may be asked, are many naturalists now-a-days not satisfied with the evidence afforded by Lord Morton's case? Why does Weismann tell us he inclines to Settegast's view "that there is no such thing as an 'infection' of this kind, and that all the instances which have been recorded and discussed critically . . . are based upon a misconception? "['The Germ-plasm,' p. 386.] Why, while the belief in infection forms an important part of the creed of breeders, did Romanes and other scientific believers in telegony think it occurred but rarely, in only one or two per cent, of cases? When Lord Morton's case is considered critically the evidence in support of infection is found to consist chiefly (1) in the colt and filly presenting quagga- or, to be more accurate, zebra-like markings; and (2) in the filly having, according to Lord Morton, a quagga-like mane while the colt had a mane like the quagga hybrid. Weismann, following Settegast and others, if asked to account for the stripes on the colt and filly, would reply by saying they were due to reversion or atavism. This answer I need hardly point out is based on the assumption that there is such a thing as reversion, and on the further assumption that the ancestors of the horse of to-day were even more striped than the recently extinguished quagga (Fig. 13). Granting reversion, and granting also that the various breeds of horses have descended from profusely striped ancestors, Weismann's answer would be quite legitimate. But some biologists, who are not prepared to allow that reversion of this kind ever occurs, would not admit the relevancy of Weismann's answer. If not adopting the "infection" explanation they would probably say the stripes on the colt and filly were an instance of excessive or discontinuous variation - their resemblance to ancestral markings being a mere accidental coincidence.

Fig. 13. - Lord Morton's Quagga (after a drawing by Agasse).
Fig. 14. - Lord Morton's Quagga Hybrid (from Agasse's drawing, Royal College of Surgeons Museum, London.).
Although it is impossible to prove that reversion occurs in horses, as it admittedly does in pigeons, I believe that if the markings on Sir Gore Ouseley's "colts" were not due to the dam having been influenced in some way by the quagga they resulted from reversion. I prefer the reversion explanation, because it seems to me to be simpler and more in accordance with established facts. I have frequently seen numerous zebra- or quagga-like stripes not only in foals but also in aged horses, stripes on the face as well as on the shoulder, neck, body, and legs. Hence the possibility of reversion may in the meantime be taken for granted, and the further question asked. Were the markings on the "colts" sufficient in themselves to prove that the Arabian mare had been infected by the quagga? I think it must be admitted that the stripes taken by them- selves are barely sufficient to prove infection. If the paintings by Agasse (in the Royal College of Surgeons' Museum, Lincoln's Inn Fields, London) are accurate, Lord Morton's quagga-hybrid (Fig. 14) was but feebly striped. Agasse shows three distinct but short stripes across the left shoulder, four faint and still shorter stripes on the neck, ten cross-stripes on the fore-leg - six of them between the knee and the elbow - and nine cross-markings on the hind leg, five of which lie above the hock. When I first saw Agasse's drawing of the quagga-hybrid I was surprised at the meagreness of the striping. In all my hybrids there are more stripes than in their zebra sire, and the general, though not universal, result of crossing zebra mares with asses and horses has been the production of abundantly striped offspring. Further, although there were no cross bars (Fig. 18) on the legs of the quagga, [Although, as far as I am aware, there are no stripes on the legs of the quaggas preserved in museums, I understand quagga foals bad sometimes striped legs.] there were more stripes on the legs of the hybrid than elsewhere. Hence in the case of the hybrid at least we seem to have certain evidence of reversion - reversion to the ancestors of the horse or the quagga, or as Mr. Galton would perhaps put it, regression to the ancestral mid-parent. ['Natural Inheritance,' London, 1889.] I have had in my possession horses with more stripes than are represented by Agasse in Lord Morton's quagga-hybrid, and hence it would be difficult to prove that this hybrid was not indebted to its ancestors on the female side of the house for all the stripes it possessed.

Fig. 15. - Filly bred by Sir Gore Ouseley from Lord Morton's Mare. (from Agasse's drawing, Royal College of Surgeons Museum, London.).
In the filly - the second foal of Lord Morton's mare - faint stripes are shown by Agasse across the fore-leg between the elbow and knee, and across the hind leg above and below the hock. On the neck there are seven stripes, and there are eight stripes across the body behind the shoulder-stripe, which vary considerably in length (Fig. 15). There are thus quite twice the number of stripes on the neck and body of the filly that are shown in the hybrid, and, unlike the quagga (Fig. 13), the filly has stripes on both the fore and hind limbs. In the colt Agasse represents a broad shoulder-baud which looks as if it consisted of two or more stripes. In front of this band there are several (eight or nine) stripes on the neck, while behind the shoulder-stripe there are others extending nearly to the loins - twelve quite distinct and two doubtful. I noted also five faint markings across the fore-leg above the knee, and two equally obscure markings across the hind leg above the hock. In the fourth foal of Lord Morton's mare (which, like the second and third, had for his sire Sir Gore Ouseley's black Arabian horse) I detected in Agasse's sketch eight stripes in all, three very faint stripes in front of, and four behind, the shoulder-stripe.
Weismann points out in reference to the stripes on the subsequent foals of Lord Morton's mare that similar stripes are "not very uncommon on purely bred foals, and ordinarily disappear as the animal grows older." I have seen eleven pairs of stripes on a Shetland foal, and Darwin, as already stated, bred a foal marked by numerous narrow stripes, some of them as far back as the croup. Nevertheless I am not sure that Weismann, in saying that stripes such as Agasse shows in the two-year-old colt are "not very uncommon on purely bred foals," gives a sufficient and final answer. While stripes at birth may be accounted for in some cases by saying they are simple relics of ancestral markings, it does not necessarily follow that they may be thus accounted for in all cases; and notwithstanding all the criticisms that have been made, it is quite possible that had Lord Morton's mare not borne a hybrid foal to a quagga her offspring to the black Arabian horse might have been absolutely devoid of stripes even as foals.
Darwin was abundantly familiar with the fact that stripes now and again appear on the shoulder, legs, neck, and face in horses in all parts of the world and of various breeds and colours. Hence I imagine that, on coming to the conclusion that the quagga had affected the character of the subsequent offspring begot by the black Arabian horse, he was not a little influenced by Lord Morton's description of the mane in the colt and filly. Undoubtedly stripes together with an erect or nearly erect mane would practically prove infection had taken place. This leads me to consider the evidence afforded by the mane in the Ouseley colts. Lord Morton, referring to the mane, says, " That of the filly is short, stiff, and stands upright, and Sir Gore Ouseley's stud-groom alleged that it never was otherwise; that the mane of the colt is long, but so stiff, as to arch upwards and to hang clear of the sides of the neck, in which circumstance it resembles the hybrid. This is the more remarkable as the manes of the Arabian breed hang lank, and closer to the neck than those of most others."
When some days ago I re-examined the oil-paintings by Agasse of Lord Morton's mare, her quagga-hybrid, and her three foals to Sir Gore Ouseley's black Arab horse, I specially noticed, as was previously pointed out by Settegast, that the mane of the filly (Fig. 15) was not represented as short, stiff, and erect, while that of the colt was not shown as arching upwards.* It is well known that the mane of the horse is easily altered. In one of my Arabs (Benazrek) the mane hangs lank and close to the neck, but in another (once a polo pony) the mane six months ago was short, stiff, and upright, while now that it is longer it arches upwards free from the neck; some months hence it will doubtless hang lank and close to the neck. Sir Gore Ouseley's stud-groom alleged the filly's mane had always been short, stiff, and upright; but this, it seems to me, is proving too much, and the point at issue is far too important to rest on an allegation of this kind.
[* I failed to ascertain the date of the paintings, but from the third pure- bred foal forming one of the series it may, I think, be taken for granted that the sketches were made during the summer or autumn of 1S21, i.e. about a year after Lord Morton made his communication to the Royal Society. Lord Morton's mare, it may be mentioned, missed having a foal in 1820. See Sir E. Home's ' Conip. Anat.,' vol. iii, p. IS, 1823. ]
The necessity of repeating the Morton-Ouseley experi- ment with a well-striped zebra became still more evident when I read in 1895 Mr. (afterwards Sir Everett) Millais' 'Two Problems of Reproduction,' ["Our Dogs" Publishing Company, Manchester, 1895] in which he states: "I may further adduce the fact that in a breeding experience of nearly thirty years' standing, during which I have made all sorts of experiments with pure-bred dams and wild sires, and returned them afterwards to pure sires of their own breeds, I have never seen a case of telegony, nor has my breeding stock suffered. I may further adduce the fact that I have made over fifty experiments for Professor Romanes to induce a case of telegony in a variety of animals - dogs, ducks, hens, pigeons, &c., - and I have hopelessly failed, as has every single experimenter who has tried to produce the phenomenon." [The late Sir Everett Millais, like Romanes, believed that telegony was possible, but he thought it was "exceedingly rare, and therefore abnormal" (loc. cit., p. 20)]
THE HYBRID "ROMULUS" AND HIS SIRE AND DAM.
Having made the necessary arrangements, I purchased in the spring of 1895 two female zebras, a number of mares of different sizes, colours, and breeds, an Arab horse, and later, a handsome Burchell's zebra (Equus hurchelli) . [This zebra might be placed in Mr. Pocock's sub-species Chapmani. (See Pocock's paper on " the Species of Zebras," ' Ann. and Mag. of Nat. Hist,.,' July, 1897); and Mr. Prazak's 'Wild Horses of Africa,' MS., The Library, University of Edinburgh.] For months no progress was made, and for a time my attempt to cross mares with a zebra stallion seemed destined to fail. In course of time, however, some of the difficulties were overcome, and on August 12th, 1896, a zebra-horse-hybrid made its appearance. Since then four other zebra hybrids have arrived, and more are expected next summer (1898) - all by the same sire. A photograph of the first hybrid (Romulus) with his dam (Mulatto) was reproduced in the Field of September 19th, 1896, and a short description of the sire, dam, and hybrid was published in The Veterinarian for November of the same year.

Fig. 16. - Mulatto.
The Dam, Mulatto. - Mulatto (lent for the experiments by Lord Arthur Cecil) belongs to a Western Highland breed, is nearly black in colour, and just under 13 hands in height (Fig. 16). This breed has long been under observation, and, except in 1848, no fresh blood has been recently introduced into the Island of Rum section of the group, to which Mulatto's sire belonged. [* The well-bred black West Highland ponies are or were found chiefly on the Inner and Outer Hebrides, which lie to the west and north-west of the mainland of Scotland - the survivors of the Island of Rum herd Lord Arthur Cecil had removed some time ago to Kent] Hence these ponies may be looked upon as forming a remarkably pure breed, and Mulatto may be considered an excellent substitute for Lord Morton's all but pure Arabian chestnut mare. [Mulatto, by producing an abundantly striped hybrid foal, has proved herself well fitted for the experiment from at least one point of view.] As to the Rum pony foals, I have learned from Lord Arthur Cecil that they are at birth frequently mouse-coloured with a faint dorsal stripe. After the first year the "colts" are dark brown or nearly black, with in most cases hazel- coloured eyes, a thick long mane and forelock, a long heavy tail, chestnuts on both fore and hind limbs, but little hair at the fetlock joints. There is no record of these foals having been marked with shoulder-stripes, though there has been occasionally for a time a cloudy patch over the shoulders; neither have stripes been noticed on the legs, neck, or body. In support of the belief of the late Marquis of Salisbury (who owned the Island of Rum) that these ponies had Eastern blood in their veins, it may be men- tioned that in various respects they approach the highest type of horses that has probably ever existed - I mean, of course, the high-caste Arabs. I need only add as to Mulatto that Romulus was her first foal, that she is now (1897) six years old, and that she is of an extremely gentle disposition, - as unlike as possible the restless, excitable, inquisitive zebra with which she was first mated. [For further particulars as to the West Highland ponies see p. 6]

Figs. 17 & 18 - Matopo.
The Sire, Matopo. - The zebra sire (Matopo) presents all the characteristic markings of a Burchell's zebra (Figs. 17 and 18), and though under thirteen hands, he is a handsome fellow, always in excellent condition, with fairly good shoulders, perfect legs and feet, and very fine action. The hind quarters are well formed, the tail is set on more as in a horse than in an ass. The warts (chestnuts), only present on the fore-legs, are large, smooth, and black. Oval in form, they measure 3 in. by 2 in., and lie as in the ass as nearly as possible at the middle of the forearm. In summer the stripes are nearly black, while the intervening spaces on the sides of the neck, body, and hind quarters are of a cream-colour; elsewhere they are nearly white. In winter brown hairs (over an inch in length on the body, but shorter on the face) grow out from the dark bands, while equally long white hairs grow out from the intermediate spaces. As summer sets in the long brown and white hairs are gradually shed, the summer coat consisting of hairs from an eighth to three eighths of an inch in length. During the summer of 1897, as in 1896, a few of the long hairs persisted on the sides of the body."
[* It is worthy of note that during the present winter (1897-98) the long brown hairs have not yet (February) appeared. This is doubtless due to the extreme mildness of the season. In its native country the winter coat is worn during our summer. This may account for some of the long brown hairs persisting during the two previous seasons well into the autumn.]
The skin is dark throughout, under the white hair the skin is of an iron grey colour, elsewhere it is nearly black owing to the pigment in the hair roots. The mane consists at each side of black and white tufts of hair, which alternate with each other; sometimes a narrow light patch an inch or two in length is introduced into the neck to admit of this alternation being fairly regular. The central part of the mane consists of black hairs measuring from five to six inches in length, except over the withers, where they are short, and between and in front of the ears, where they are from three to four inches in length; the black central hairs are always longer than the white lateral tufts, the difference in length being greatest in the front half of the mane. Both the dark and light hairs of the mane are shed annually, and the light tufts are the same colour summer and winter - not white in winter and cream-coloured in summer, like the spaces in the neck with which they are continuous. The mane extends on to the forehead four inches beyond the occipital crest, to end in hairs which project directly forwards im- mediately above a tuft of hair," quite two inches in length, that grows upwards at one side of the mane from the point where the lower frontal stripes meet. [* I shall speak of this tuft of hair as the frontal tuft; a similar but usually shorter tuft is found in the horse under the forelock. The forelock, which is sometimes considered a special growth, consists simply of the front part of the mane. ] The ears, which are wide and rounded at the tips but not very long (6-and-a-half inches measured along the upper surface), are lined with almost white hair, and have dark margins. When projected forwards the ears form with the mane three broad bands, which like the stripes melt into grey by moonlight.
The upper part of the tail (about ten inches) is transversely striped, and one third of the striped part is covered with short hair similar to that on the hind quarters. From the terminal part of the tail long dark persistent hairs grow out, which reach to within a foot of the ground. The intermediate part of the tail bears hairs which are shed annually. These hairs vary in length from two to eight inches; some are black, while others are white. In the autumn some of these hairs form a crest continuous with the dorsal band, while others project outwards to form two lateral fringes. I need only mention further, as to the colour of the hair, that though the muzzle looks black in some lights it looks grey in others. This is due to the fact that the all but black skin is only sparsely covered by very short light hairs. Above each nostril the hair is of a brownish colour. In the ill-defined "nostril patches" several of the facial stripes terminate. Between the dark brown " nostril patches " the hair is almost black; in the Somali zebra, it may be mentioned, the muzzle some- times in colour resembles that of a typical Exmoor pony (is, in fact, "mealy-coloured"), while the nostril patches are of a bright tan colour.

Fig. 19. - Matopo.
Fig. 20. - Romulus.
Fig. 21. - Somali Zebra iE. grevyi).
In Matopo, as in most of the Burchell zebras, the stripes on the forehead have a somewhat lozenge-shaped arrangement (Fig. 19). Two stripes, one starting above each eye, meet in the middle line near the end of the mane to form an acutely pointed arch - the orbital arch. Above this pointed arch are four rounded arches; the lowest ends in the frontal tuft already alluded to, the others extend into the mane. Within the orbital arch are two distinct and still more acutely pointed arches. The limbs (each being double, as is also the case with the orbital arch [From the limbs of these arches being double it may be inferred that Matopo's ancestors had at least double the number of frontal arches]) of the outer of these arches curve inwards on a level with the eyes, and rundown to end in or near the nostril patches. The limbs of the inner arch bifurcate about the level of the eyes, and also proceed towards the muzzle, one of them again forking on the way. Within the innermost acutely pointed arch are two nearly vertical stripes; both of these split where they blend with the arch above, while the right one again splits before it reaches the muzzle below. The three pointed arches and the first rounded arch meet in the middle line, and from their junction the long hairs grow out which form what I have spoken of as the frontal tuft.
I have described thus fully the stripes on the front of the face of Matopo that a comparison may be afterwards made between his acutely pointed frontal arches and the numerous rounded arches in the hybrid offspring, Romulus (Fig. 20). To admit of further comparison with the hybrids it is necessary that I should also describe at some length the stripes on the body of Matopo. Not only are no two individuals of any given species or variety of zebras alike in their marking, but in no single individual is the pattern the same on both sides.* But notwithstanding the varia in individual zebras, and the extreme cases of variation within the same species, it is possible to recognise a general plan in their marking. I consider the Somali zebra (Fig. 21) the most primitive of all the zebras. This conclusion has been arrived at mainly from a compara- tive study of the markings of zebras. I have examined a number of skins from Somaliland (one of which was a well-preserved and quite complete foetal skin), numerous Burchell's zebra skins, several skins of the mountain zebra, and the Amsterdam, Leyden, London, and Edin- burgh quaggas.
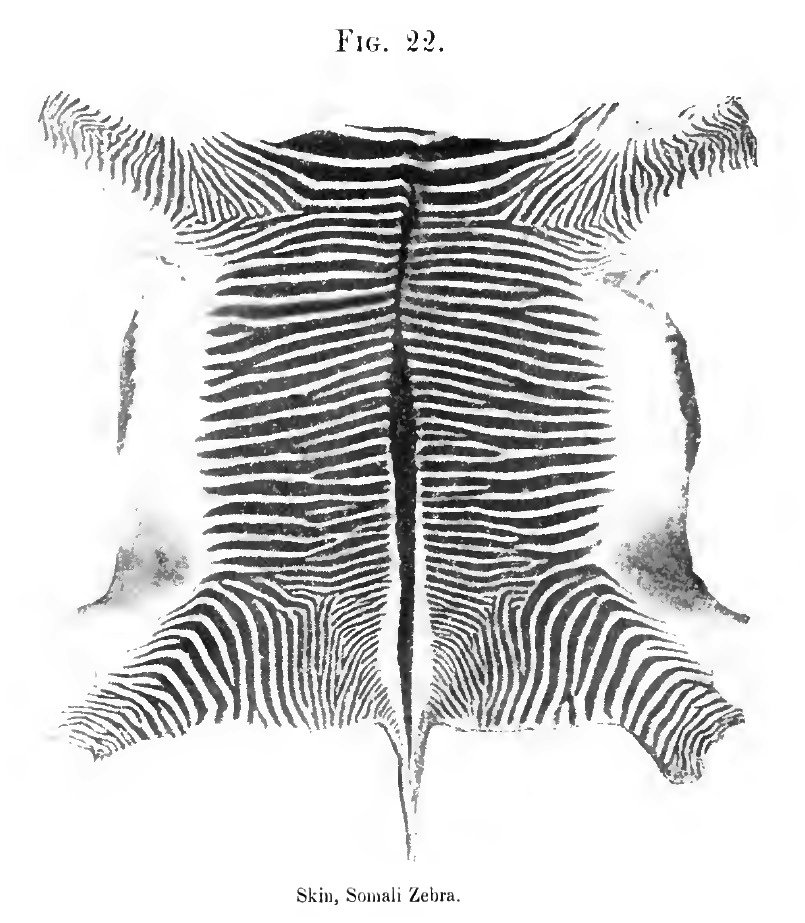
Fig 22. - Skin, Somali Zebra.
[* As far as I can learn, want of symmetry in the colouration of plants and animals is comparatively common. On the other hand, in plants and probably also in animals, a point is eventually reached in the difference of the two sides which is prejudicial to the life of the variety or species. Were not this the case, wild animals would doubtless be as asymmetrical in their colouration as our artificially protected domestic animals. While intercrossing makes for asymmetry, inbreeding seems to lead to symmetry in the markings. Mr. Prazak tells me the Craddock, an inbred race of mountain zebras, are remarkably symmetrical in their markings.]
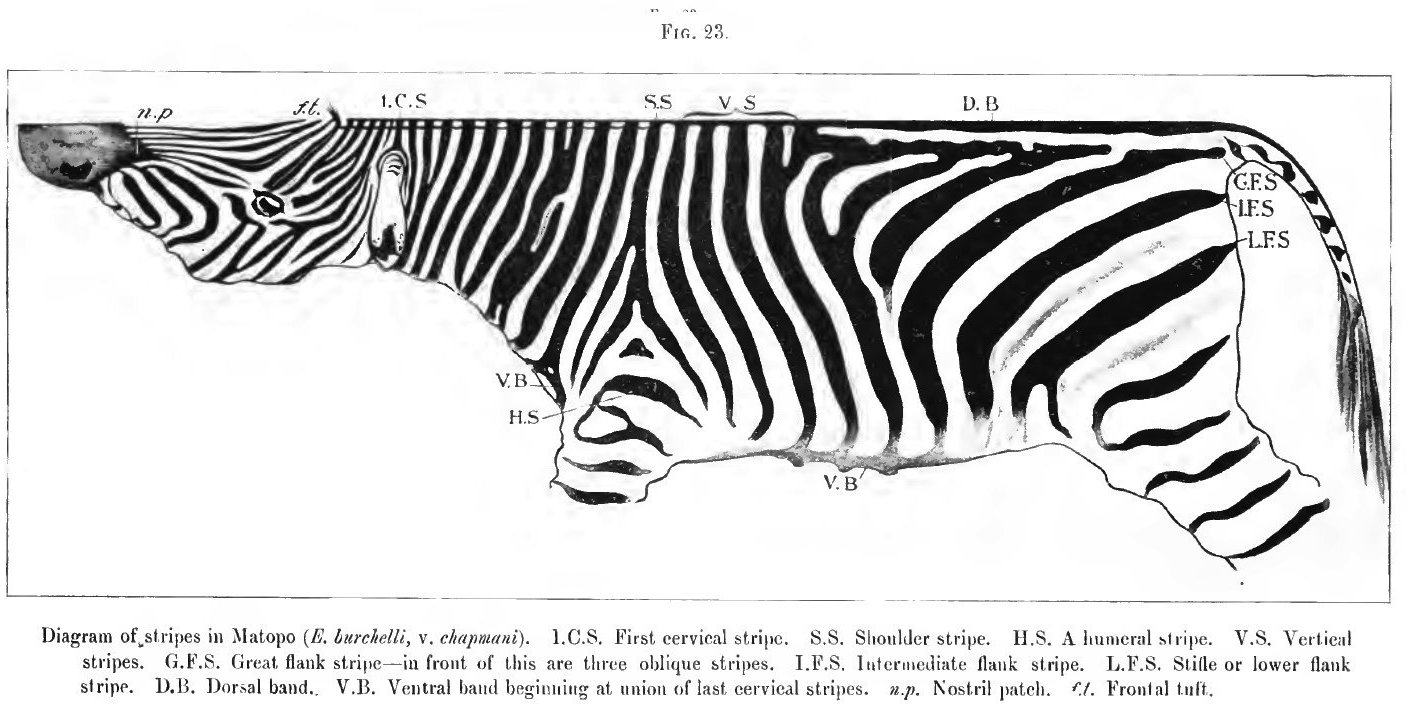
Fig 23. Diagram of stripes in Matopo.
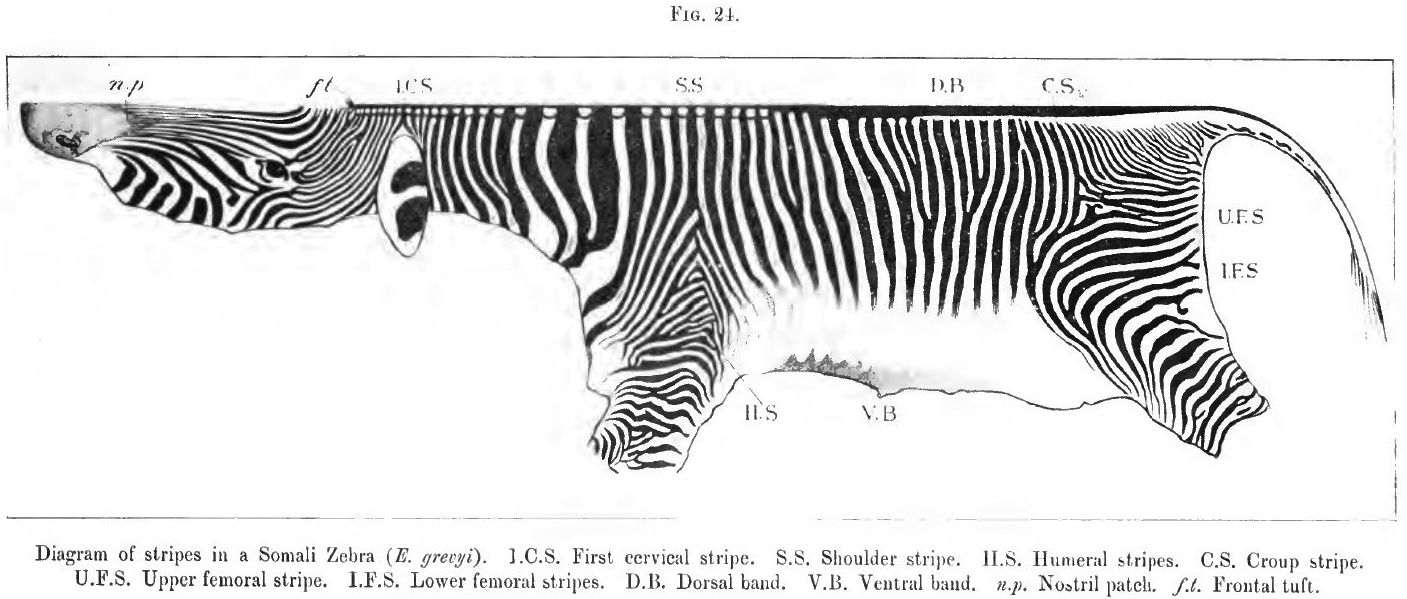
Fig 24. Diagram of stripes in a Somali zebra.
The chief difficulty in dealing with zebras is to find possible points of comparison in the marking of the different species and varieties. In all the zebras there is, however, one very distinctive stripe, viz. the stripe which typically extends downwards from the withers to bifurcate somewhere above the level of the shoulder-joint; one division proceeding forwards across the shoulder-joint, the other backwards behind the elbow. This characteristic stripe I shall speak of as the shoulder stripe (Figs. 17, 18, and 23, S.S.). [* In Matopo neither of the divisions of the shoulder stripe reach the ventral band; this band usually begins at the junction of the two stripes lying immediately in front of the shoulder stripes. Mr. Prazak, I think rightly, looks upon the shoulder stripe even when consisting of a single band as made up of two stripes (' Wild Horses of Africa ').]
Occupying the triangular space formed by the two limbs of the shoulder stripe and the transverse band at the base of the forearm, we find in the Somali zebrat some ten narrow arches (the humeral stripes) (Fig. 24, H.S.), the upper ones pointed, the lower ones rounded. [By the Somali zebra I invariably mean the Somaliland variety of Grevy's zebra, not the typical Grevy's zebra (E. grevyi, Oust.) of Shoa.] In Matopo (Fig. 23) only portions of three humeral stripes can be made out, but a Burchell's zebra filly (Fig. 25) from the Transvaal had at least seven. Between the shoulder stripe and the occipital crest there are in Matopo twelve stripes continuous with black bands in the mane (Fig. 23). In the Transvaal filly there are twelve stripes on the right side and ten on the left, but as there is a "shadow" or indistinct stripe (Fig. 25) running down the majority of the light bands in this filly, there were probably at least twenty-four cervical stripes between the occipital crest and the shoulder stripe in the ancestors of the zebras. In the Somali zebra there are sometimes fourteen cervical stripes extending into the mane, but judging by the width of some of them a considerable amount of blending has evidently taken place.

Fig. 25. - Young Burchell's Zebra with "shadow stripes."
Fig. 26. - Mountain Zebra (E. zebra).
Fig. 27. - A Typical Burchell's Zebra (E. burchelli).
A careful examination of the hind quarters of the Somali zebra reveals the fact that there is a stripe which has nearly the same relation to the hip-joint as the shoulder stripe has to the shoulder-joint. This, the croup stripe (Fig. 24, C.S.) (which typically consists of two stripes), also when single bifurcates; one limb running backwards across the hind quarters, the other forwards to unite with the first of a series of distinct stripes which arch across the hind quarters, and may be known as the femoral stripes (Fig. 24, F.S.). Between the shoulder and croup stripes in the skin figured (Fig. 24) there are twenty-five stripes running at nearly right angles to the dorsal stripe. But as four of these are more or less forked at their origin, the twenty-five probably represent at least twenty- nine vertical stripes. Hence it may be inferred that in still more primitive zebras there were numerous narrow not very well defined stripes running vertically across the body between the shoulder and croup stripes. As in the case of the shoulder stripe, several arches (modified femoral stripes. Fig. 24, U.F.S.) occupy the space between the two divisions of the croup stripe. The anterior limbs of these arches usually blend or all but blend with the first of the complete femoral stripes (Fig-. 24, I.F.S.). Between the croup stripe and the root of the tail there are at least twenty narrow stripes which arch backwards. These stripes begin some distance from the dorsal band,* and form with the corresponding- stripes of the opposite side a series of arches which diminish in size from before backwards (Fig. 22). Trans- verse markings extend across the tail to near where the long hairs of the tip spring; in some cases not a few of the caudal markings blend to form two narrow bands - one at each side of the tail.
[* In some of the Somali zebras the narrow transverse (croup) stripes are at their origin only a short distance from the dorsal band; in others a fairly wide light space intervenes between the croup stripes and this band. But when the croup stripes all but reach the dorsal band, they are for an inch or two only faintly coloured.]
It is difficult if not impossible to identify a croup stripe in the mountain zebra (E. zebra, Fig. 26), or in any of the Burchell zebras. Nevertheless I shall endeavour to show that a certain amount of resemblance exists between the stripes of the Somali and all the other zebras. In Matopo a pronounced band may be traced from the root of the tail (with the stripes of which it is in line) forwards over the external point of the ilium (haunch) across the flank, to curve downwards and inwards and finally end in the ventral band. This, which may be known as the caudal or great flank stripe (Fig. 23, G.F.S.), is present in all the Burchell skins I have examined. With the exception of the Burchell zebra proper, there are in all the Burchell varieties and sub-species I have seen two other well-marked flank stripes. The lowest of these (Fig. 23, L.F.S.) is intimately related to the stifle (knee proper), at or near which it frequently bifurcates, one division, usually slender, running down in front of the stifle, the other or main division (Fig. 18) extending forwards and inwards to- wards the ventral band. This may be known as the stifle or lower flank stripe. The other flank stripe (Fig. 23, I.F.S.), which lies between the caudal and stifle stripes, may be known as the intermediate flank stripe. [In some zebras one or more of the flank strii)es may bifurcate or unite with one of the adjacent stripes as it proceeds towards the ventral band.] In Matopo the third flank stripe stops short of the ventral band, while in the true Burchell zebra (Fig. 27) the third flank stripe is at the best feebly developed. Between the great flank stripe and the vertical stripes there are in the crawshayi zebras a number of oblique stripes, some of which reach the ventral band. In the common zebra, the stripe (Figs. 26 and 28) which forms one of the handles of the "gridiron" [The stripes across the croup and rump, together with the two great flank stripes, may be said to form a " gridiron " with two long handles.] I look upon as corresponding to the caudal or great flank stripe of Matopo; it has the same relation to the root of the tail, and is the third stripe in front of the stifle. Is it possible to detect in the Somali zebra stripes corresponding to the three flank stripes in the common and Burchell zebras? I look upon the femoral stripe, which in the Somali zebra is intimately related to the stifle, as the equivalent of one of the oblique stripes lying between the vertical and flank in the Burchell zebras. This stripe, which in the Somali zebra forms a frame for the insertion of a number of transverse stripes, in all probability also corresponds to one of the long curved handles into which is inserted the transverse base of the "gridiron" of the mountain zebra. That there is some relation between the femoral stripes in the Somali zebra and the oblique and flank stripes of the other zebras will be more evident when the croup and flank and femoral stripes in zebra hybrids are taken into consideration.
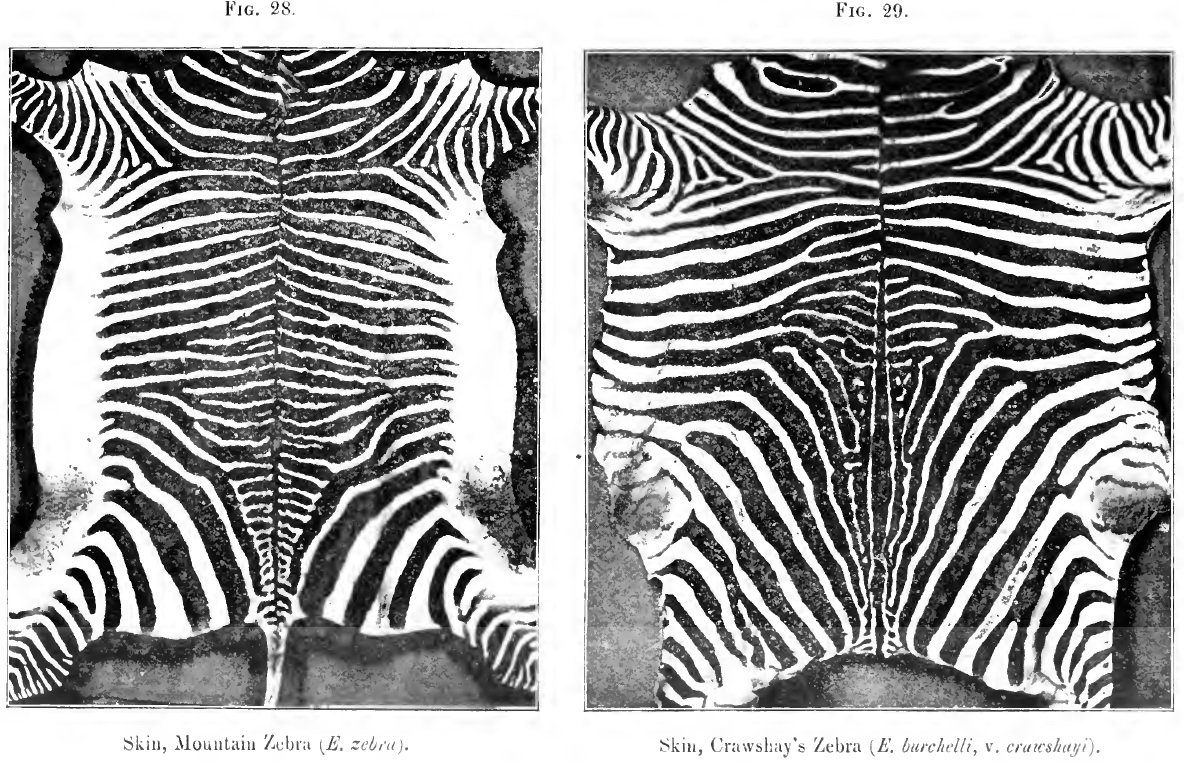
28. Skin of a mountain zebra
29. Skin of a Crawshay's zebra
From the stripes in the mountain and Crawshay zebras (Figs. 28 and 29) it is comparatively easy to derive the varied patterns which obtain in the Chapman and Burchell zebras. Between the lower part of the stripe which forms the frame of the "gridiron" in the mountain zebra (Fig. 28), and the shoulder stripe, there are in the skins I have seen from nine to ten nearly vertical stripes crossing the sides, the majority of which in some cases reach, or all but reach, the ventral band (Fig. 30); while in the Somali zebra at the same level there are from eighteen to twenty stripes, some of which after a short break also reach the ventral band.*
[* Pocock, in referring to the common zebra (E. zebra), says, "Except for the longitudinal ventral band the belly is white, the flank stripes stopping short of the belly as in the quagga, Burchell's zebra, and Grevy's zebra." In the common zebra I saw last March in the Amsterdam Zoological Gardens, stripes extended right across the belly, and in the foal (now, 1898, in the Zoological Gardens, London) the long broad ventral fringe of hair was partially striped like the mane. This striation of the ventral fringe (which has not, as far as I am aware, been ever described) is quite evident in a photograph (Fig. 30) for which I am indebted to the Director, Dr. C. Kerbert. In the Somali zebra some of the vertical stripes may reach the ventral band. As they cross the belly they get lighter and lighter, until they are almost invisible; but when within two or three inches of the ventral band they become nearly as distinct as they are on the sides of the body.]
As the number of stripes in zebras is evidently sometimes reduced either by two or more blending, or by alternate stripes first forming shadow stripes and then disappearing, the nine to ten vertical body stripes in the common zebra may be equivalent to the eighteen to twenty stripes in the Somali zebra. In the Burchell zebras, when the shadow stripes (Fig. 25j are included, there are often eleven vertical stripes* between the shoulder stripe and the great flank stripe, i.e. the stripe corresponding to one of the handles of the gridiron in the common zebra.[* In Matopo four vertical stripes reach the ventral band (Fig. 23), while in the Transvaal filly (Fig. 25) three stripes on one side and two on the other reach the ventral band.]

Fig. 30. - Mountain Zebra and Foal (Amsterdam Zoological Gardens).
Hence the only essential difference between the Burchell and the common zebra is that in the latter the great flank stripe has numerous cross-bars extending from it towards the dorsal band. This gap between the mountain and Burchell's zebra is (as far as the markings are concerned) bridged over by members of Crawshay's group of zebras. Hence, as already indicated, it appears (1) that the arrangement of the stripes over the flank and hind quarters in Matopo has been arrived at by the femoral stripes of the Somali zebra arching upwards and forwards so as to entirely obliterate the numerous narrow stripes across the croup and rump; (2) that a less complete migration of the femoral stripes would result in the plan which prevails in the Crawshay type of zebras (Fig. 29); and (3) that such a "gridiron" as exists in the mountain zebra would almost at once result from the shunting upwards of the posterior end of the first complete femoral stripe in the Somali zebra. In other words it appears that the plan of marking in the common zebra might be easily derived by a modification of the stripes in the Somali zebra, and that by further modifications in the same direction the various patterns presented by the stripes in the Crawshayi, Chajpmani, and Burchelli types of zebras might also be obtained. I do not wish it to be inferred that the Burchell zebras have been derived from the common zebras, but simply that the ancestors of the Burchell zebras once upon a time more or less resembled in their markings the common zebra of to-day, and that their still more remote ancestors probably resembled in their markings the Somali zebra.
It need only be mentioned further as to Matopo, that below the knee and hock the stripes are faint; that while the front part of the pastern is white, the back part with the exception of a mesially placed white triangular space is decorated with very narrow dark brown stripes; that there are no hairs at the fetlock; that the chestnut (wart) on the fore-leg - the only one present - while relatively much larger than in the horse, is smooth and not raised above the general level of the skin; that the hairs (vibrissae) about the muzzle are longer and more numerous than in well-bred horses; and that only the five vertical stripes behind the shoulder stripe blend Avith the dorsal band, which, as in many zebras, expands slightly as it crosses the croup.

Fig. 31. - Romulus (seven days old) and his dam, Mulatto.
Fig. 32. - Romulus: twenty-seven days old.
Fig. 33. - Romulus: twenty-seven days old.
Romulus. - In Romulus, and in my other hybrid foals, there are numerous stripes, but, as it happens, the plan of the striping- is in all of them quite unlike that of their sire Matopo. This will be at once evident if the photographs of Romulus (Figs. 31, 32, and 33) are compared with those of his sire (Figs. 17 and 18) . In Romulus (a year old on the 12th of August, 1897) the stripes on the forehead (Fig. 33) form a series of rounded instead of, as in his sire, pointed arches (Fig. 19). In two of this year's hybrids there are similar arches across the forehead, but in the others the pattern of the frontal stripes is somewhat different. Romulus, in having rounded instead of pointed frontal arches, agrees with the Somali and Crawshay's zebras (Figs. 34 and 35). I have photographs showing the markings on the face in five Somali zebras. No two are alike, but in no case do the arches form acute angles, as in Matopo (Fig. 19). In all the hybrids the stripes on the forehead are more numerous and consequently narrower than in the Somali zebra. [In Romulus there are fourteen narrow dark brown curved frontal lines, with equally narrow pale lines between them.]
In the striping of the neck, body, hind quarters, and legs Romulus also resembles the Somali zebra far more than his sire. In the case of Matopo there are twelve stripes which extend into the mane between the occipital crest and the shoulder stripe; in Romulus there are twenty-four stripes in the same position. In the Somali zebra there are, on an average, fourteen cervical stripes; but evidently in this zebra, as already mentioned, a certain amount of blending of the cervical stripes has taken place. In Gray's figure of Burchell's zebra, shadow stripes are represented in front of as well as behind the shoulder stripe. ['Zool. Journ.,' pl. iv, July, 1825.] Notwithstanding Gray's figure I have heard doubts expressed as to shadow stripes being present on the neck of zebras.
[The danger in dealing with zebras is coming to conclusions from the examination of any given collection of skins or specimens. Conclusions arrived at to-day often stand in need of altering to-morrow, so great and remarkable is the range of variation in the stripes, more especially in what may be known as the Burchell group of zebras. Though familiar enough with the variations in the markings of our domestic animals - which for many generations have been removed from the influence of natural selection - we are hardly prepared to find a wide range of variation in the colour and marking of zebras living in a wild state in comparatively limited areas. That in the case of the Burchell group of zebras (i.e. the Crawshayi, Chapmani, and Burchelli zebras) there is a wide range of variation in the arrangement, colour, extent, and relations of the stripes, in the mane and tail, and in the form of the head, neck, and hoofs, will be abundantly evident if an attempt is made to classify them. A satisfactory classification will not be possible until a large number of skins are available, the district to which each skin belongs being in each case specified.]

34. Somali zebra, face markings
35. Crawshay's zebra, face markings
There is, however, no room for doubt on this point. In my two zebra mares there were distinct shadow stripes on the neck, and I have before me a photograph of a zebra in the Dublin Zoological Gardens showing four very distinct shadow stripes on the left side of the neck. [I am indebted to Professor D. J. Cunningham, P. U.S., of Dublin, for this photograph.] In one of my mares, which I understand was captured in the Transvaal when a few weeks old, there were ten shadow stripes on the left side of the neck (Fig. 25). These shadow stripes, together with the twelve almost black stripes, made twenty-two cervical stripes in all. On the right side there were, in this mare, eight shadow stripes, which with ten ordinary stripes made in all eighteen - a difference in the two sides of four stripes. Evidently even the Burchell zebras have had at one time a considerable number of distinct cervical stripes; and Romulus in having twenty-four cervical stripes, if he does not reproduce an ancestral plan, in all probability approaches nearer to it than his sire Matopo. Between the shoulder stripe and the root of the tail there are, as already indicated, in the Somali zebra over forty stripes running outwards at each side at nearly right angles to the dorsal band. In Romulus forty-three stripes or portions of stripes may be made out between the shoulder stripe and the root of the tail: in the common zebra, notwithstanding its gridiron arrangement, there are under thirty stripes. The first ten of these stripes in Romulus seem to take the place of the five broad nearly vertical stripes in Matopo. The stripes beyond the tenth are at first narrow and indistinct, while over the loins and croup they were at birth mainly made up of rows of spots. Now that Romulus is a year old many of the spots over the rump have united to form somewhat zigzag stripes. [This, I think, proves that at least the transverse stripes over the croup and rump, in the Somali and common zebras, have been formed by the running together of rows of spots.] The majority of the stripes over the rump run at nearly right angles to the dorsal band, but as the root of the tail is reached some of them bend backwards, and thus very closely agree with the narrow recurved stripes in the Somali zebra. In two of this year's hybrids the rump is less marked than in Romulus, while in one there has been from the first all but complete narrow wavy lines in place of the spots originally present in Romulus.
Although the arches within the two limbs of the shoulder stripe are in Romulus almost as indistinct as were the markings in this region in the quagga, they in a favourable light are seen to be seven in number, i.e. as numerous as in the Transvaal filly (Fig. 25) . Below the elbow, however, the transverse markings are relatively broader and more pronounced than in Matopo. On the outside of the leg eighteen stripes can now be counted, while there are seventeen on the inside. The spaces between the stripes are of a dark mouse-colour from the elbow to immediately below the knee, but they get gradually darker as the hoof is reached, until it becomes almost impossible to detect them. The inside of the leg is lighter than the outside, the upper part of the forearm being of a drab colour on its inner aspect. The oblique and flank stripes are not very well defined in Romulus. There is, however, a stripe which seems to correspond to the great flank stripe in his sire, though in its position it more closely agrees with the first complete femoral stripe in the Somali zebra. This, the third in front of the stifle, extends upwards to arch across the upper end of the flank feather, to lose itself amongst the spots over the hind quarters. [I propose to call the "swirl" or hair-shed that usually extends in the horse between the stifle and the point of the ilium the flank feather. A corresponding but shorter swirl, which I have occasionally seen extending upwards from near the elbow, may be known as the elbow feather.]
The second (intermediate) flank stripe runs obliquely across the feather, expanding considerably near where the feather begins. After a considerable interruption this stripe, as Fig. 32 shows, again appears as a narrow tapering band running parallel to a more distinct stifle stripe lying at a lower level. Now (1898) by the blending of spots the inter- mediate stripe is more complete on the left side than at birth, while on the right side a number of spots have recently blended to form a continuation of the upper flank stripe. It thus appears that longitudinal as well as transverse stripes may originate from the blending of spots. The stifle stripe appears to begin at the bottom of the flank feather. It then runs for some distance parallel to the middle flank stripe, and after a short interruption crosses the hind quarters as a well-marked band, to end some distance below the root of the tail. The three flank stripes, while differing in their position from the flank stripes in Matopo, agree fairly closely with stripes in the Somali zebra.
In front of the flank stripes are three oblique stripes which, beginning on a level with the stifle, run upwards parallel with the flank stripes to lose themselves amongst the spots and indistinct narrow bands over the loins and croup. Similar oblique stripes occur in some of the crawshayi type of zebras. In the space bounded in front by the last vertical stripe, above by the dorsal band and below by portions of the oblique and flank stripes, are numerous nearly transverse stripes and rows of spots. The part of this space in front of the croup is mainly occupied by in- distinct narrow stripes, over the croup proper there are interrupted more or less zigzag stripes, behind these numerous spots. Round the root of the tail an attempt has been made to form a series of semicircular lines (Fig. 11) such as occur in the Somali zebra (Fig. 24). The "gridiron" of Romulus thus agrees more closely with the markings over the loins, croup, and hind quarters of the Somali (E. grevyi) than with those over the corresponding parts of the mountain zebra (E. zebra).
In describing the flank stripes in the hybrid I mentioned they crossed the flank feather. In horses this swirl or feather usually extends from near the level of the stifle to near the point of the ilium or haunch-bone, while in asses it is short, and usually nearly circular in form. In Matopo there is no swirl formed by the hair in the region of the flank, nevertheless there is a fairly long feather in my zebra hybrids. In Romulus it begins about four inches above the level of the stifle, and extends relatively as high as in the horse. [The flank swirl being quite small or absent in the ass, it is not surprising to find that in a zebra-ass hybrid the feather or swirl is wanting.] Between the lower flank stripe and the hock stripe - the stripe which typically bifurcates as it crosses the hock - there are ten distinct transverse bands, between some of which fairly distinct shadow stripes may be seen. Altogether it is possible to identify thirteen stripes above the hock when the three flank stripes are included. Below the hock the stripes are now (1898) rather obscure, owing to the darkening of the intermediate spaces. It is, however, possible to make out some thirteen stripes on the outside of the leg below the hock. The inside of the leg above the hock is of a drab or mouse-colour. Within six inches of the stifle there is a straight stripe running across the leg, and between this line and the hock eight other stripes are present. While the outside of the hind shank in Romulus has as many stripes as in his sire, the inside is distinctly less striped; but, on the other hand, the pastern and fetlock are much darker.
In Romulus there are only chestnuts on the fore-legs, but in Remus - a bay Irish mare's hybrid - there are chestnuts on both fore and hind legs, and in a Clydesdale's hybrid there is a small chestnut on one hind leg. In all my hybrids, as in their sire, there are ergots at the fetlocks, but only in two of the hybrids is there a distinct tuft of hair at the fetlock. The mane in Romulus agrees with that of his sire, except that many of the central hairs reach, before they are shed, a length of eight or nine inches, and the difference between the colour of the lateral tufts is not pronounced. In Matopo the mane extends as far as the frontal tuft, but in Romulus there is a gap of about half an inch between the tuft and the mane. From the terminal part of the tail, which is relatively longer than in the horse, a bunch of long persistent hairs grow; but from the greater part of the tail the hair was shed during last winter, and developed again during the spring.
In the Burchell zebras the eyes are less prominent than in the horse. This is partly due to a slight insinking of the eyeball and the absence of a prominent supra-orbital ridge. In the hybrids the eyes may be said to unite the characters of both parents. The lower eyelid instead of inclining inwards as in the zebra is nearly vertical in position, while the supra-orbital ridge is more prominent. The upper eyelashes are, however, peculiar, and the upper eyelid never seems to be raised as high as in the horse - only the lower half of the eyeball is as a rule exposed . The upper eyelashes in all the hybrids, but especially in Romulus and the very zebra-like Irish mare's hybrid Remus, are long and curved, so that even when the upper lid is quite raised the long hairs arch downwards in front of the eye. In my Arabs and in Matopo the upper eyelashes are comparatively short, and when the lid is raised they project almost directly outwards. In Matopo under the lower eyelid the skin is coloured so as to resemble a fringe; similar marks are found in zebra hybrids, especially in zebra-ass hybrids. The nostrils in Romulus and the other hybrids are zebra-like, narrow, short, and almost oval in form, and not deeply notched as in the horse at the upper angle; and the rounded outer margin of the nostril projects freely outwards as in zebras and asses.
It thus appears that hybrids obtained by crossing different breeds of mares with a Burchell's zebra stallion differ in many respects from both their respective parents, and yet, except in a few points, they can hardly be said to occupy an intermediate position, to resemble the ideal "mid-parent" that presumably combines the characters of the two immediate ancestors. This is true not only of their colour, but also of their make, habits, and disposition. They differ from their respective dams in being elaborately striped, they differ from their common sire in the number and disposition of the stripes. But what is perhaps most remarkable is that, while differing from all the Burchell zebras in the number and arrangement of their stripes, all the hybrids either closely agree with the Somali zebra or present what I consider a simpler and more primitive arrangement, as, e.g., having twenty-four bands in the neck, and in some cases having rows of spots instead of transverse stripes in the region of the rump. The hybrids may be said to take after their sire in their nostrils, their hoofs, and to a less extent their chestnuts, and in the males having two teats. [Foetal colts have frequently, if not always, distinct rudiments of teats up to about the sixth month. Relatively large teats are usually present in the jackass and in male mules.] They may be said to be intermediate in their make as a whole, but especially in their ears, eyes, neck, withers, and hind quarters, and in their mane and tail. In having a somewhat long head they agree with the Somali zebra, but in having long curved eye-lashes they are, as far as I am aware, unlike the majority of both horses and zebras. In the dorsal band being bounded along its whole length on each side by a narrow yellow band, they differ from their sire, and again approach the Somali zebra, in which there is a light space at each side of the dorsal band for more than half its length.
WERE THE ANCESTORS OF THE HORSE STRIPED?
The fact that all the five hybrids differ in their markings very decidedly from their sire, while they in many respects suggest the Somali zebra, demands an explanation. Darwin seems to have been not only impressed with the difference between hybrids and their immediate ancestors, but also with their striking resemblance to known or supposed remote ancestors. [Mr. Francis Galton has long recognised that hybrids differ from their immediate ancestors. He quite recently pointed out that "the offspring of two diverse parents do not necessarily assume an intermediate form." -Nature, Oct. 21st, 1S97, p. 599.] He accounted for this resemblance by saying crossing had led to reversion. As the question of reversion is of the utmost importance in considering supposed cases of telegony, it will be necessary to deal with it at some length. There is, however, a prior if not equally important question, viz.. Have all our breeds of horses - Arabs, Exmoors, Clydesdales, &c. - descended from striped ancestors, from hog-maned ancestors, elaborately decorated with bands on the face, neck, body, and legs, after the fashion of one or other of the "gaily painted zebras?" Evidently it is desirable to consider this question before dealing with the reversion hypothesis.
What evidence is there in favour of the view so generally held that the ancestors of the horse were striped? Over four thousand separate books have been published on subjects appertaining to the horse, and though many of these works devote a chapter to the horse of antiquity, I believe I am right in saying that they afford no evidence of a horse striped after the manner of a zebra or quagga having existed during the historic period. What of the prehistoric period? Every one knows that the palaeolithic cave-dwellers, instead of devoting much time to polishing their implements, after the fashion of their neolithic successors, spent some of their leisure in delineating on bone, slate, and other permanent materials the beasts of the chase - amongst others the mammoth, reindeer, and horse. Now although these palaeolithic men lived many thousands of years ago, they were evidently accurate observers. A sketch, e.g., of a reindeer fight on a piece of slate is most spirited, while that of a mammoth on a piece of ivory (found in the Madelaine cave) is strikingly accurate. But what of the horses? That the Troglodyte artist was abundantly familiar with the horse will be evident when I mention that near Solutre, in the neighbourhood of Macon, to the north of Lyons, there are the remains of thousands of horses which had undoubtedly served as food during a considerable period for a fairly large settlement of primeval men. The bones of the horse are mingled with - to mention only ungulates - those of the mammoth, Canadian deer, primeval ox, and the saiga-antelope. The majority of the bones are broken, the long ones having been split for the sake of the marrow. Do any of the sketches, made it may be thirty thousand years ago, throw any light on the striping of the horse?
Some think they do. They certainly give evidence of remarkable powers of observation; the legs, e.g., are almost as accurately placed as in a Muybridge photograph. Yet, notwithstanding a number of suggestive lines, these primeval sketches can hardly be said to help us in answering the question, Were the ancestors of our various breeds of horses striped? Though nothing definite can be learned from either historic or prehistoric records, there may be other available evidence. Every one knows that stripes are not by any means uncommon, more especially in dun-coloured Norwegian and Kattiawar ponies. Are these markings vestiges of a striped coat, as complete, it may be, as that of the quagga, or are they feeble attempts on the part of some recent horses to mimic the richly decorated zebra? Before attempting to answer this important and, it must be confessed, difficult question, let us see to what extent some of the horses of to-day are striped, and the nature of the pattern. First-hand evidence in matters of this kind is most valuable. I shall, therefore, in dealing with the striping of the horse, refer almost exclusively to stripes present on the horses I have specially examined during the last three years.
Head Stripes. - In a sand-coloured (yellow dun) Norwegian pony, with black mane and tail, which has been in my possession for over a year, there are stripes on the face, neck, body, and legs. [I may mention that this is a typical Norwegian pony, the dam of which I have no hesitation in saying never had the opportunity of even seeing a zebra.] On the forehead there are two all but complete frontal arches and portions of five others (Fig. 36) . Being of a reddish-brown colour these stripes are easily seen when the forelock is thrown back. The upper-most (orbital) arch ends in the frontal tuft, but instead of forming an acutely pointed arch, as in Matopo, it forms a somewhat rounded arch, as in the Amsterdam quagga and in one of my zebra hybrids. The fragments of the other arches are most distinct in the middle of the forehead. ln having seven more or less complete frontal arches this pony differs from my Burchell zebra, in which there are only three distinct arches in the corresponding position. When the incomplete arches are "restored" a pattern is formed which is almost intermediate between that of the Amsterdam quagga and the Somali zebra. From within the lowest arch several obscure lines, such as occur in zebra-ass hybrids, can be traced along the face.

Fig. 36. Norwegian Pony.
The stripes doubtless originally ended in or near a mealy-coloured muzzle, such as we find to-day in typical Exmoor ponies and in some Somali zebras. I have seen only two ponies with stripes on the face, and I imagine facial stripes are extremely rare. Darwin as a youth seems to have been devoted to horses. When at Cambridge he had a passion for shooting and hunting, and when that failed riding across country. This may partly account for his great interest in horses in after years, for his systematically collecting information about horses from all parts of the world, that he might, if possible, come to some conclusion as to the original colour and markings of their less remote ancestors; Apparently Darwin only once noticed stripes on the face. This was in a cob-like fallow-dun having a conspicuous spinal stripe and its front legs well barred. Darwin, however, learned from Colonel Poole that Kattiawar horses had often " stripes on the cheeks and sides of the nose." ['Animals and Plants,' vol. i, p. 61.] I have only seen faint indications of stripes on the sides of the face, but from what I have seen I have no hesitation in saying that were a sketch made showing all the stripes of which fragments have been observed on the face of the horse during the present generation, the sketch would closely resemble the head of one of my hybrids and less closely the Amsterdam quagga. As stripes on the forehead in the form of acutely pointed or rounded arches are, as far as I am aware, only found in the horse family, and as stripes are evidently being gradually eliminated in horses, most will agree with me that the facial stripes in the Norwegian pony are not new creations, but vestiges of ancestral markings. [Were they new creations they would, if the usually accepted view as to the origin of stripes is correct, in all probability consist of rows of spots in process of blending, instead of narrow, more or less distinct lines. Further, the "blaze," so common in some breeds - whether in the form of a lozenge or of a long white patch extending to the muzzle, - is easiest accounted for by supposing the face of the ancestral horse resembled that of the quagga or one of the zebras.]
As to the ears, I may mention I have seen two light dun-coloured ponies - one from Shetland and one from Norway - with the tips for quite half an inch almost white. In both cases a dark broad band extended across beneath the light tip. In the Norwegian with the striped face there are only a few light hairs at the tip of the ears, but immediately below there is a broad black band, and an indistinct band near the base. Had the tip been lighter in colour the ear of this pony would have not a little resembled in its decoration that of my Burchell zebra.
Neck Stripes. - The Norwegian pony has only a short shoulder stripe, but in an old bay Highland cob in my possession a year ago the shoulder stripe was nearly a foot in length. Sometimes the shoulder stripe bifurcates some distance above the level of the shoulder- joint, and thus suggests not so much the zebra as the quagga and zebra hybrids. In the Norwegian pony there are a number of ill-defined stripes in front of the shoulder stripe, and in another light dun-coloured pony there are ten cervical stripes. As these ten stripes only extend about halfway along the neck, and as stripes are sometimes present immediately behind the ears, there may have been quite twenty stripes in the ancestors. As a rule the neck stripes are short and indistinct, but in the Highland cob one extended quite two thirds across the neck; and in some cases I have seen three or four cervical stripes nearly as well defined as in the zebras. In one case I observed several stripes extending into the mane.
Body Stripes. - In the Norwegian with the partially tattooed forehead there are only three short stripes on the body behind the shoulder stripe, but there were six in the Highland cob. Mr. Darwin, in discussing striping in horses, says, "Stripes on the body, not to mention those on the legs, are extremely rare - I speak after having long-attended to the subject - with horses of all kinds in Europe, and are unknown in the case of Arabians." ['Animals and Plants,' vol. i, p. 435.] I have, however, before me a photograph of a mouse-dun Norwegian pony showing vestiges of stripes nearly as far back as the loins - as far back as in the Gore Ouseley filly (Fig. 15). But it is not only important to prove that there were stripes on the body, but also to determine if possible their number and direction, and to ascertain whether they most agreed in their disposition with those of the Burchell, the mountain, or the Somali zebra. In the yellow-dun Norwegian there is an extremely well developed dorsal band, as distinct and as broad as it crosses the croup as in my Burchell zebra. Darwin says, "I have never heard of either shoulder or leg stripes without the spinal stripe." [Animals and Plants,' vol. i, p. 63.] The spinal stripe or dorsal band, in fact, occurs more or less constantly in all the species and varieties of the horse family. This band and the leg stripes are so common and persistent that it looks as if they had either been established long before the other stirpes or retained long after the stripes on the face, neck, and body had all but disappeared.
When one looks from a height on the dorsal band of the Norwegian pony, the edges are seen to give off short processes - rudiments of developing stripes or vestiges of dwindling ones, such as are seen in some of the quaggas and in zebra-ass hybrids. Once, and once only, have I seen as many as five distinct stripes extending from the dorsal band across the back. Evidently vertical stripes on the body, as Darwin observed, are extremely rare - almost as rare as on the face.
But although well-defined stripes are seldom seen on the trunk, obscure delicate lines may often be detected running- down the sides of the body and across the flanks in horses of all breeds and colours. These lines I first noticed last May in a dark yellow-dun Canadian mare - they were gradually revealed as the winter coat was shed. [This and another mare Mr. Stewart, cab proprietor, Dean Bridge, Edinburgh, was good enough to lend me for some weeks last spring.] These lines, which vary in width from an eighth to a quarter of an inch, though in most cases difficult to see, are sometimes (when lighter in colour than the intervening spaces) quite distinct. Usually they are most evident when the coat is "staring" slightly. At times, as Mr. Alexander, R.S.A., said, they remind one of the lines on a roll of bacon - the lines produced by the string with which it is for a time encircled. [I am not aware of any reference to these lines by recent writers Mr. Alexander alone, of all those wise about horses I have consulted, had seen them; he has even tried to represent them in some of his pictures.]
In the yellow-dun Canadian mare there are seventeen lines between the elbow and the stifle, separated by spaces about two inches in width. The four most posterior arch upwards in front of the stifle, and then curve backwards across the flank feather. The lines which cross the lower part of the feather are quite evident in the majority of horses; they are particularly distinct in two of my chestnut mares, and in an almost white Arab mare. All but continuous with these "feather" lines I have seen ill-defined markings extending right across the hind quarters of two bay horses of the Shire breed. In front of the recurved lines there are in the Canadian mare thirteen others, which occupy a nearly vertical position. Some of them incline forwards as they pass behind the elbow, to end in what might be described as an indistinct ventral band; while others bend slightly backwards. In Mulatto relatively broad vertical lines are especially distinct. [One of the recognised differences between recent horses and their allies, the zebras, is the absence in the horse of a ventral band; but in the Canadian mare an indistinct band, about a foot in length, extends along the breast-bone.] Beginning some distance behind the shoulder-blade, within seven or eight inches of the spinal ridge, they run down the sides of the body towards the ventral surface. At a distance of twelve yards seven lines can be easily recognised, the second forking about the level of the shoulder-joint, after the manner of the body stripes in some zebras and zebra hybrids. In some cases I have seen lines running obliquely upwards above the level of the flank feather. These delicate and faint flank markings (which are distinct enough in a yearling chestnut filly to admit of their being photographed) are especially interesting, because they closely correspond to narrow stripes in a similar position in my zebra hybrids. [This filly belongs to the Walltower Stud Farm. To the owners (Messrs. Cairns) I am indebted for many favours during the last two years.]
Have these various lines any significance? Are they due to rows of hair lying at an unusual angle, or to furrows in the skin, or are they the last remnants of ancestral stripes, or of the spaces between stripes? [I may mention that the stripes have no relation to the ribs or to the course of nerves or blood-vessels in recent zebras, but they may have run parallel to nerves or blood-vessels in the ancestors of the zebras. Some of them, however, correspond closely to wrinkles which occasionally appear on the skin; but this does not hold for the neck stripes.] Somewhat similar but more prominent lines occur in Indian cattle, the ancestors of which were probably striped, and also in mules, and in some of my zebra hybrids. Taking into consideration that these lines in their arrangement agree more or less accurately with the stripes in the mountain zebra, and with the fragments of stripes in Norwegian and other ponies, I am inclined to the view that though not necessarily vestiges of stripes they indicate where the ancestral stripes occurred. All we know as to the origin of stripes is against the supposition that the lines are the rudiments or forerunners of stripes by-and- by to be evolved.
Let us now inquire if stripes occur across the croup (sacral region) and rump - the region lying between the croup and the root of the tail. Darwin describes stripes in this region in a foal of his own breeding. After mentioning that almost the whole body was marked with very narrow obscure stripes, he says the "stripes were distinct on the hind quarters where they diverged from the spine and pointed a little forwards." Darwin does not seem to have observed stripes across the croup in a full-grown horse. Such stripes are, however, said to occur even in horses having a large proportion of Eastern blood. I have before me a drawing of a well-bred Austrian mare which, in addition to shoulder stripes and stripes on the face, neck, and legs, and across the root of the tail, had three stripes across the hind quarters. One of these runs obliquely backwards and outwards across the croup, while the two others extend across the rump. In their direction these markings probably differed from the stripes on the "hind quarters" of Darwin's colt, from the rump stripes in the common zebra, and, as will appear later, from markings across the croup in Mulatto's second foal. As stripes in the horse have long counted for little or nothing in the struggle for existence, these oblique stripes may not bear any relation to ancestral markings. Although I have never seen transverse markings across the croup and rump of horses - markings such as characterise the common zebra - it does not follow they never exist. It is quite conceivable that in some breeds of horses they are comparatively common. It may be here mentioned that indistinct markings were sometimes present on the hind quarters of the quagga; that irregularly arranged spots are very common over the hind quarters of zebra-ass hybrids, and that, as already mentioned, there were at birth spots over the croup and rump of Romulus. These facts point to the croup and rump stripes having been acquired comparatively late in the zebras, and to their having begun to vanish almost before they were firmly established - before they were sufficiently burned in to persist when neglected by natural selection. On the other hand I have frequently seen a cloudy patch over the croup and rump of bay horses which, seems to indicate that in the horse stripes persisted long over the hind quarters.
Leg Stripes. - I have only now to refer to the striping of the legs. So common are stripes on the legs of horses, that whatever is thought as to the presence of stripes on the face or body, no one seems to doubt that the legs of the ancestors of the horse were provided with numerous transverse markings. Leg stripes, more especially in the vicinity of the knee (wrist) and hock (ankle), seem to be nearly as old as the dorsal band. In some yellow and mouse duns I have seen stripes on the fore limb from the fetlock up to within a short distance of the elbow, and on the hind limb a considerable distance above the hock. As in the zebras, the stripes are sometimes distinct, at other times they have more or less run together. In some zebras the leg below the knee is almost black; perhaps the much admired "black points" of the favourite bay have resulted from the fusion of ancestral stripes on the lower part of the legs. From a consideration of all the stripes I have seen on horses, I have come to the conclusion that the ancestral horse was in its markings nearly intermediate between the common and the Somali zebras. In Romulus we may have a fairly accurate restoration of the stripes in the ancestors of the Burchell zebras and, though less likely, also of some of the markings in the ancestral horse.
Having indicated to what extent recent horses may be striped, the mane and tail may next be considered. The attempt to arrive at any conclusion as to the mane and tail of the ancestral horse may seem quite hopeless. Endeavouring to settle whether the mane was long and lank, or short and upright, and whether the tail could only boast of long hairs at the tip, in say the horse of the Pliocene epoch, may seem to some attempting the impossible. We sometimes hear of palaeozoologists reconstructing an entire skeleton from a single bone- it may be from a single tooth. I think we have the equivalent of at least a fossil tooth in the markings on the face of the Norwegian pony. These markings, distinct enough now (October), were strikingly evident in May, after the winter coat was shed. Nevertheless they were unobserved for months, for the simple reason that they were hidden under a massive forelock. I may again say that I look upon the incomplete stripes on this pony's forehead as relics of a time when the horse had a striped face, not as rudiments of stripes in process of development. Stripes are almost invariably preceded by spots not very regular in their arrangement - spots that if joined in one way give longitudinal stripes, if in another transverse. The evolution of stripes such as we find in zebras must have occupied count- less ages, and their maintenance in a complete form must have involved the suppression of not a few interesting variations. It is almost inconceivable that nature would decorate the brow of a horse after the pattern of a zebra if the work when completed would never be seen. Hence we may take one of two things for granted; (1) either that the process of tattooing is proceeding now, or (2) that the brow markings are vestiges of stripes which existed in, and were of use to, the ancestors, before a great frontal bunch of hair or forelock made its appearance. That the last supposition is most in keeping with what we know of nature's methods will be at once admitted. But if the forelock was originally so short that the frontal stripes were visible, the mane must also have been short; for the forelock is not a special growth, it is nothing more nor less than the most anterior part of the mane, that part of the mane which lies between and in front of the ears. In other words, the period in the ancestral history of the horse characterised by a complete series of facial stripes was, it may be inferred (if the stripes were of any use), also characterised by a mane consisting of short nearly upright hairs which were annually shed.
In support of the view that the primeval horse had a short upright mane, it may be mentioned that in colts the mane is as a rule upright for several months, and that it sometimes eventually falls to the right side, sometimes to the left, sometimes partly to one side in front and to the other side further back. Further, when in Norwegian dun ponies the long dark central hairs are cut, the finer light- coloured hairs at each side naturally assume an erect attitude. Of still more importance is the fact that the wild Tarpan, to which some of our recent horses are doubtless related, had an erect or nearly erect mane, the central hairs of which were presumably annually shed. [The mane is said to be still sometimes upright in the domestic horse, and it is represented as erect in palaeolithic etchings.] The retention of the hair of the mane until it reached a considerable length may have been found useful, especially when lying down, to horses living in cold, damp, northern areas, where stripes were of little or no value.
In all probability the hair of the mane lengthened and became more persistent as the hair of the body increased in length and thickness. That the coat became thicker and longer as the horse extended its range to mingle with the reindeer and the other tundra fauna may be taken for granted. While the long hairs of the mane and tail were probably first acquired by natural selection, they may have been retained and lengthened by artificial selection. The Arab, probably evolved in a cold area, has retained the long mane and tail; but the hairs, like those of the body generally, are apt to be fine and silky. Occasionally, according to Azara, horses were born in Paraguay with hair like that on the head of a negro. In such horses the mane and tail were short - they were, in fact, curiously correlated to the hair on the body. ['Animals and Plants,' vol. i, p. 56.] This further supports the view that the long mane was acquired after the horse left its ancestral home - perhaps long after it migrated from the New into the Old World.
There is in the horse family for some reason or other always an intimate relation between the condition of the mane and that of the tail. In the recent horse the mane consists of long persistent hairs (sometimes the mane has been over six feet in length), and the tail also from within two or three inches of the base to the apex of the dock consists of persistent hairs, those growing from the tip being sometimes long enough to reach the ground. In the zebra, as already stated, the mane, as a rule, consists of short, upright, deciduous hairs, while the tail, though having long persistent hairs at its tip, has deciduous (never very long) hairs elsewhere. In zebra-horse hybrids the hair in the mane, though sometimes double the length of that in the zebra, seems to be shed annually; and though there are longish hairs up to the base of the tail, only the long hairs springing from and near the tip persist. [I have seen a zebra in which the mane, as in zebra hybrids and in the ass of Somaliland, was long enough to fall slightly backwards and to one side; a further development might result in a horse-like mane.]
That the tail in the ancestral horse resembled that of the zebra of to-day may also be gathered from a study of its development. In the embryo the tail is at the outset not only quite hairless, but much longer than the hind limbs. By-and-by the limbs grow so rapidly that the tail hardly reaches as far as the hocks. At about the eighth month of foetal life the tail (with the exception of its tip) is covered with quite short hairs; the tip has already longish hairs. In fact, the history of the development of the tail, as far as made out, seems to indicate that in the remote ancestors of the horse there were long hairs at the apex only, as in the Somali zebra; that in less remote ancestors less long but deciduous hairs extended up to within a short distance (six or seven inches) of the root, as in the Burchell zebras; and that eventually, as the hair began to persist in the mane, persistent hairs appeared up to, or almost up to, the base of the tail, as in our recent horses.
What answer shall we now give to the question: Have our horses descended from striped ancestors? Taking into consideration the facts I have mentioned, and especially (1) that there are more or less striped horses in all parts of the Old World; [See ' Animals and Plants,' vol. i, p. 61.] (2) that in Mexico and various other parts of the New World the descendants of horses introduced by the Spaniards are frequently dun-coloured and provided with dorsal and leg stripes; [Nature, March 11th, 1897.] (3) that foals are often partially striped, one bred by Darwin being marked all over with obscure narrow stripes; and (4) that mules (hybrids between the jackass and mare) have frequently leg stripes as well as spinal and shoulder stripes - taking these and other facts into consideration, the conclusion may be hazarded that the ancestors of all our horses were striped, and that the stripes were arranged more after the plan in the common than in that of the Burchell zebra. If we agree with Darwin "that not even a stripe of colour appears from what is commonly called chance," ['Origin of Species,' p. 129.] I fail to see what other conclusion could be arrived at.
REVERSION IN THE EQUIDAE.
Having given reasons for the belief that the ancestors of the horse were striped - striped more or less after the pattern of the mountain zebra, - the important question may now be asked: Do horses without so much as a dorsal band sometimes bring forth offspring with a considerable number of stripes? and the still more important question: Are the stripes in such offspring due to reversion? It is "widely believed that stripes are not very uncommon at birth even in Arabian foals. I may add some recent evidence in support of this belief. Two years ago I bred a foal from a black Shetland pony, the sire being also black and of the same strain as the dam. [The sire was Wallace, a champion Shetland pony.] The foal, which was of a reddish dun colour, presented at birth the following stripes: - (1) a dark dorsal band which extended a short distance into the tail; this band widened as it reached the croup, and again, as in many zebras, contracted as it approached the root of the tail; [When the foal was two months old, this band consisted mainly of erect reddish hairs (such as I have seen in zebra and zebra hybrid foals), from which one is tempted to infer that in the remote ancestors of the Equidae the mane extended right along the whole length of the back.] (2) a shoulder stripe over six inches in length; (3) one distinct and five indistinct stripes on each side of the neck immediately in front of the shoulder stripe, and three on the body behind the shoulder stripe; and (4) a number of very obscure stripes on the legs. In this foal the mane was black and at first nearly erect, while the tail was of a dark grey colour. When six weeks old the foal began to shed its coat, and when two months old the markings had completely disappeared from the legs and the sides of the body, and they were less distinct on the m ck. When a year old this colt was of a dark bay colour, but only retained the dorsal band and two stripes across each shoulder. It may be well to point out that both the dam and the sire may have been striped at birth, and that had they been bay instead of black, some of the stripes might have been visible throughout life." [Is it possible that we owe our black horses to the blending of dark ancestral stripes?]
Mr. Wilfrid Scawen Blunt has been good enough to send me some particulars as to his 1897 crop of Arab foals. Mr. Blunt has this year ten foals from pure Arab mares, all the foals having as their sire Mesaoud, a bright chestnut Arab with four white legs and a blaze. Of seven bay mares, five had bay foals while two had chestnut foals. Of the five bay foals two had a dorsal band. One of the mares which, like the sire, is a chestnut had a chestnut foal with a dorsal band. Of the two remaining mares one is grey, the other white; the grey mare had a bay foal, the white a chestnut, showing distinct zebra markings inside the forearms, less distinct markings above the hocks, and a hardly perceptible line down the spine. Mr. Blunt informs me that the dam of this striped colt is absolutely pure white (and, like the sire, of the most perfect Arab type); that he has noticed that stripes occur most frequently in foals from white parents, but that the stripes do not outlast the second year, or at longest the third. The desert Arabs seem to dislike dun-coloured horses, and, probably owing to the duns being weeded out, stripes are comparatively rare in desert-bred high-caste Arabians. Nevertheless four (i.e. 40 %) of Mr. Bhunt's 1897 Arab foals had a dorsal band, and one in addition stripes on both the fore and hind legs. It is especially interesting to note that all the ten foals were either bay or chestnut. The foals of grey horses are never born grey; with few exceptions, foals at birth belong to the red or mouse dun series of colours - are bay, chestnut, or brown. If both parents are of the same colour and strain and inbred, the foals may closely resemble the parents even if they are black or cream-coloured or piebald.
We can only guess as to the colour of the remote ancestor of the horse, but nearly all who have made a special study of the subject have come to the conclusion that the less remote ancestors were dun-coloured. But it is hardly sufficient to say the ancestors were dun-coloured, for in Norway four shades of dun are recognised, which include nearly every colour from white to black. There are (1) white duns (white and light creams) with white mane and tail; (2) yellow duns with black mane and tail, including creams and light bays; (3) elk duns, frequently approaching in hue bays, chestnuts, and browns; and (4) mouse duns, some of which are nearly black. After a full consideration of the subject, I am inclined to believe the body colour of the striped ancestral horse of the temperate regions was mainly of a yellowish brown colour. As the descendants extended their range the ground colour would change, a sand colour probably prevailing in desert areas, a reddish dun in the vicinity of forests, a mouse dun in the far north, a light tint near the tropics, and in the uplands a grey or ash tint.
From various experiments made, it seems to me that black, white, grey, and cream-coloured horses, even if striped, would have little chance of surviving long enough in a wild state in the Northern Hemisphere to form black and other conspicuously coloured breeds, and that the most serviceable colour for northern horses, i.e. the most protective colour, would be an indescribable shade of dun approaching a sand colour on the one hand, and a dull lustreless light bay on the other. The least conspicuous pony in my possession is a typical Exmoor with the characteristic mealy-coloured muzzle, black points, and the body generally of a yellowish brown or dirty bay shade, the belly and upper parts of the inside of the legs being of a uniform dirty white.
Black, flea-bitten, cream, and skewbald ponies are in most circumstances extremely conspicuous, while the mouse and dark yellow duns, the bay and brown ponies, and the ash-grey or khaki-coloured asses are in some lights often hard to see. From a yellowish brown tint the various shades of yellow, red, and mouse duns might be easily derived; and from a combination of these shades the formation of bays, chestnuts, and browns would be only a matter of time or of artificial selection.
In the foals of all shades of Arab, English, Norwegian, and Iceland horses, being with few exceptions at birth of a bay or chestnut tint, and frequently provided with a dorsal band and with leg stripes which ordinarily disappear, we have, it seems to me, evidence of reversion. For a remarkable instance of the transitory striping of a foal we are indebted to Darwin. In a chapter bearing on the colour and stripes of the horse he says: "The most interesting case which I have met with occurred in a colt of my own breeding. A bay mare (descended from a dark brown Flemish mare by a light grey Turcoman horse) was put to Hercules, a thoroughbred dark bay, whose sire (Kingston) and dam were both bays. The colt ultimately turned out brown, but when only a fortnight old it was a dirty bay, shaded with mouse grey, and in parts with a yellowish tint; it had only a trace of the spinal stripe, with a few obscure transverse bars on the legs, but almost the whole body was marked with very narrow dark stripes, in most parts so obscure as to be visible only in certain lights, like the stripes which may be seen on black kittens. These stripes were distinct on the hind quarters, where they diverged from the spine and pointed a little forwards; many of them as they diverged became a little branched, exactly in the same manner as in some zebrine species. The stripes were plainest on the forehead between the ears, where they formed a set of pointed arches one under the other, decreasing in size downwards towards the muzzle; exactly similar marks may be seen on the forehead of the quagga and Burchell's zebra. [A figure showing the extent or arrangement of the stripes on this colt would have been invaluable.] When this foal was two or three months old all the stripes entirely disappeared." [' Animals and Plants under Domestication,' vol. i, 2nd edit., p. 60.] As no mention is made as to the existence of stripes in the immediate ancestors of this striped foal, it may be taken for granted that they were whole-coloured. But one at least of the parents may have been striped at birth. On reading the description of Darwin's colt with the all but invisible stripes it naturally occurs to one that many foals may be striped at birth, but owing to the lines being so subtle they escape the notice of the ordinary observer; and further, that foals not showing stripes at birth may have passed through a striped stage before birth.
A year ago Mr. Selous was good enough to tell me of some of his experiences with zebras in South Africa. I was particularly anxious to learn about the colouration of unborn zebras. The origin of stripes has long interested and puzzled naturalists. It has been suggested that transverse stripes are preceded by longitudinal bands, the longitudinal bands undergoing a kind of segmentation, the segments being afterwards joined together to form transverse stripes. Fortunately Mr. Selous had seen an unborn zebra, which though distinctly striped looked as if it was quite destitute of hair. But as the zebra's skin is of a nearly uniform dark colour throughout, there can be no doubt that with the help of a lens hair rudiments at least would have been detected in this foetus. From the information Mr. Selous was able to give, and from an account of a foetal zebra given by Sparman, I am satisfied that the transverse stripes of zebras are not preceded by longitudinal bands. In young zebras the stripes have often a wavy, almost a zigzag appearance. Hence the transverse stripes, though not derived from longitudinal bands, may in some cases be represented for a time by spots, which by being somewhat out of line give rise when welded together to more or less wavy stripes. Are the horse foals, which are whole- coloured at birth, marked like a zebra before birth? or, Is the striped phase in the ancestral history frequently entirely skipped by an abbreviation in the development process? As I have in my possession a series of foetal horses ranging from three weeks to over ten months, I am able to state definitely that all horse-foals do not pass through a striped stage. On the other hand, though a dorsal band and leg stripes are present in one of the parents, the offspring may be quite devoid of stripes. The most remarkable instance I have come across of stripes being few and obscure in the offspring, though plentiful in one of the parents, is a zebra hybrid (Fig. 37) bred by Lady Meux. [To Lady Meux, of Theobald's Park, Hertfordshire, I am indebted for the permission to have photographs taken of her three extremely interesting horse male, zebra female hybrids.]
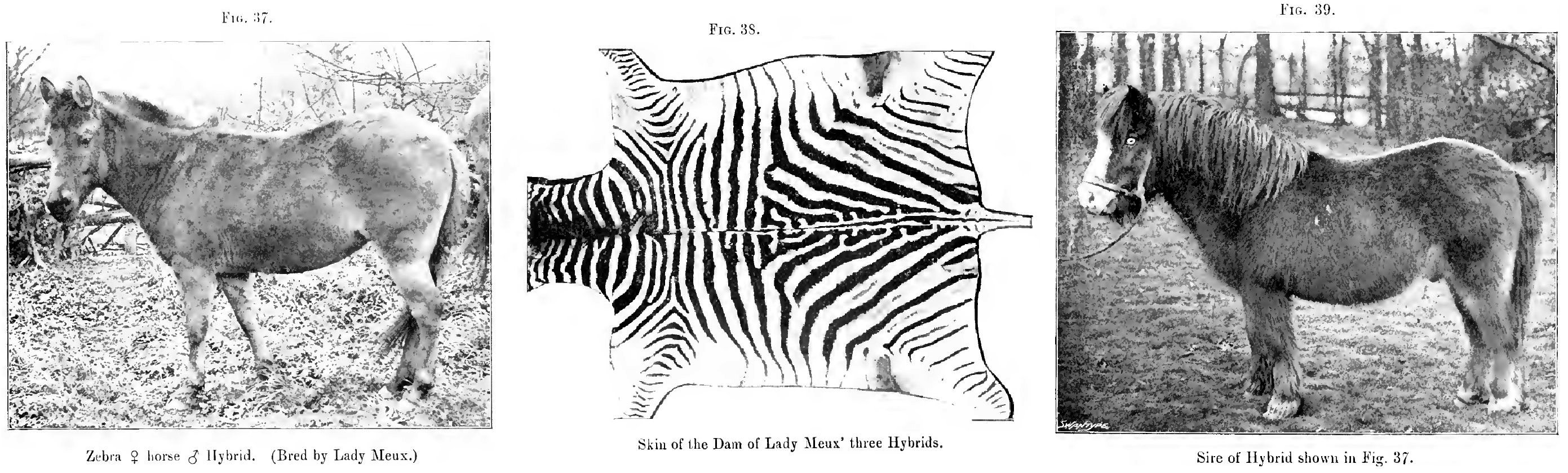
Fig. 37. - Zebra female, horse male Hybrid. (Bred by Lady Meux.)
Fig. 38. - Skin of the Dam of Lady Meux' three Hybrids.
Fig. 39. - Sire of Hybrid shown in Fig. 37.
The dam in this case was a Burchell's zebra mare (the skin, Fig. 38, of which has been preserved), the sire a small wall-eyed light bay pony (Fig. 39). The zebra mare had previously borne two foals striped after the manner of my zebra-mare hybrids. The sire of one of these foals, I understand, was a bay pony, of the other a brown pony. Why the third foal was obscurely striped is an interesting problem, which need not be here considered. The point of interest is that a profusely striped dam belonging to an ancient wild species should have produced an obscurely striped foal to a cross-bred pony, the ancestors of which have long been under domestication.
It thus appears that parents devoid of stripes may have more or less striped offspring; that all foals do not pass through a striped stage during development; that foals may lose the majority of their stripes soon after birth, and that though one of the parents is striped all over, the offspring may only possess faint indications of zebra-like markings.
Having seen that foals have often stripes at birth which afterwards disappear, the question of so vital importance in connection with telegony may now be asked: Are the stripes often found on foals due to reversion or atavism? A direct answer to this question is of course impossible, but an answer sufficiently convincing may be arrived at by the deductive method.
It seems to be admitted that all the breeds of pigeons have descended from the blue rock pigeon (Columba livia), and it has long been known that when distinct varieties of pigeons are crossed, the young sometimes very closely resemble blue, rocks. Mr. Darwin refers to a pigeon descended from a red spot and a white fantail on the one side, and two black barbs on the other. This pigeon was hardly distinguishable in its colouration from a wild rock pigeon. ['Animals and Plants,' vol. ii, p. 14.] Darwin accounted for the occasional resemblance of cross-bred pigeons to the rock pigeon, by saying they had undergone reversion. To quote from Weismann: "Darwin was the first to point out that in cross-breeding, either of species or of mere varieties, characters not infrequently appear in the descendants which were not present in the parents; in some of which cases, indeed, it can be proved, and in others shown to be very probable, that they have been derived from remote ancestors." ['The Germ-plasm,' p. 316.]
Reversion has, many think, been proved to occur in pigeons, and it is said to be very probable in the horse family. In Chile, horses living under conditions not unlike those enjoyed by their ancestors in Andalusia, are said to have remained unaltered; whereas in the eastern parts of South America the horses some years ago were chiefly dun-coloured with large coarse heads. Darwin thought this difference might also be due to reversion. ['Animals and Plants,' vol. i, p. 54; vol. ii, p. 6.] Mr. T. D. A. Cockerell recently pointed out that "in New Mexico one frequently sees small, usually pale yellowish-brown horses, with extremely well- marked leg stripes." He believes the small horses '"are descendants of the horses which ran wild in former years over New Mexico," and that there can be little doubt "that. they represent an atavistic variety." [Nature, March 11th, 1897, p. 439, I am indebted to Mr. Cockerell for a photograph showing the leg stripes in one of these New Mexican ponies.]
But hybrids between members of the horse family afford still stronger evidence of reversion. Lord Morton's quagga hybrid, though but faintly striped on the body, had distinct transverse bars on the legs, i.e. stripes which were neither present in the dam nor in the quagga sire; and many mules have often leg stripes, and occasionally shoulder stripes in addition to a dorsal band. Some months ago I saw in the South of France a light- coloured mule with the legs striped as distinctly as in the ass of Somaliland. Frequently, though there are numerous stripes on the mule, neither the sire nor dam has either leg or face stripes. I have already described at some length the plan of the stripes in my yearling hybrid. From this description it appears that Romulus differs in a most pronounced manner in the number and arrangement of his stripes, not only from his sire but from all the varieties of the Burchell group of zebras. But if one may judge from the markings on the Norwegian with the striped face, Romulus also differs in his markings from the less remote ancestors of his dam.
To recapitulate: Romulus, in having numerous narrow arches across the forehead, differs greatly from his sire, and in a less degree from the Norwegian pony; in having twenty-four cervical stripes he differs from his sire, but closely agrees with the Transvaal zebra filly when the shadow stripes are included; in having over forty stripes between the shoulder stripe and the root of the tail he is surprisingly unlike his sire, in which there are only five distinct transverse body stripes; and in having at birth well-defined spots over the croup and rump, he differed not only from his sire but from all the other zebras; and if in Mr. Darwin's striped colt we had a restoration of a lost chapter in the history of the horse, Romulus also differed in this respect from the less remote ancestors of his dam. As already pointed out, Romulus in the brow markings and in the numerous stripes running at right angles to the dorsal band, more or less accurately agrees with the Somali zebra, with which he also nearly agrees in the relative length of the head, the markings within the two divisions of the shoulder stripe, and over the rump and hind quarters. Further, at birth his muzzle resembled in colour that of the Somaliland zebra. These resemblances to the Somaliland zebra become doubly interesting when it is remembered that there are excellent reasons for believing this zebra to be more primitive than all the other living zebras. They, in fact, all but prove that in the striping of Romulus we have a remarkable instance of reversion. Additional evidence of reversion we seem to have in the intermediate spaces in the hybrids, being at first of a yellowish-brown colour, and the mane being nearly upright, and in the tail having only persistent hairs at the tip. But even stronger evidence in this direction is afforded by the spots which at birth were scattered over the hind quarters. I am satisfied these spots have no relation whatever to the dappling- so often seen in recent horses. Dappling, I believe, has been acquired since the ancestral stripes were all but lost. When dappling co-exists with more or less distinct stripes it is at once evident from the relation of the dappling to the stripes that the one has not been derived from the other. The pigment which formerly produced stripes (now no longer counting in the struggle for existence, and hence neglected) has in recent times been as it were left uncontrolled, with the result that it frequently gives rise to ever-varying and quite meaningless dappling, or to large equally meaningless blotches.
We look upon the spots on the young lion as ancestral relics, as indicating that lions (which are now in their colouring best adapted for a life in the desert or the open veldt) were at one time adapted for a jungle or forest life. In the same way, from the spots which frequently appear in young pigs and afterwards unite to form bands, we argue that the remote ancestors of the pigs were spotted, while the less remote ancestors were characterised by longitudinal bands. There are various cogent reasons for believing that stripes were in many cases preceded by spots, and we know that animals frequently during development and growth repeat more or less accurately certain phases of their ancestral history. The fact that the spots over the hind quarters of Romulus have, since his birth, to a considerable extent blended to form stripes, which almost agree line for line and curve for curve with the croup and rump stripes in the Somali zebra, seems to me to be only capable of one explanation. In the spots we have restored by what we term reversion or atavism a lost stage in the evolution of the stripes over the hind quarters in the Equidae; and by the gradual blending of the majority of these spots during the first year of the hybrid's existence, we have had a practical demonstration of how some of the stripes on the Equidae were originally formed. If the difference between the markings in Romulus and his sire are not due to reversion - to the restoration in a modified form of one of the phases through which, it may be, both horses and zebras passed during their evolution, how are they to be accounted for? The only other feasible explanation that occurs to me is, that in the numerous stripes across the sides, and in the spots over the hind quarters of Romulus, we have an instance of an abrupt form of variation. On this assumption, the resemblance of Romulus in his markings to a Somaliland zebra must be considered a mere coincidence.
The evolution of stripes in the Equidae doubtless occupied many centuries. Stripes imply at the outset a concentration of pigment in certain definite areas, and later the disposition of the pigment to form hardly appreciable ill-defined bands, or it may be to first form irregularly arranged spots, which afterwards unite to form wavy bands, the edges of which are later smoothed. But stripes in the zebras also imply a lightening of the intermediate spaces and other intricate and complex changes. By examining the hair of Romulus one seems to get a hint of how the striping has been brought about. The hairs making up many of the dark bands are at their inner ends of the same or almost the same colour as the inner ends of the hairs in the light intermediate spaces. At present (October) the hairs in the region of the shoulder are quite two inches in length. If some hairs are taken from the shoulder stripe and others from one of the light-coloured spaces near the shoulder-stripe, the inner ends of both samples are seen to be of a yellowish-grey or ash colour. But while the inner ends agree, the outer two thirds or so distinctly differ. The hairs from the dark stripes for quite two thirds of their length are dark brown or almost black, while the hairs from the light space are for quite two thirds of their length of a pale ash colour, while the remaining third - the outer or distal part - is of a light brown colour. [the difference in the colour of the hairs is more pronounced in summer.] It is doubtless possible that the result of crossing extreme forms may be the formation of new and unheard of varieties - may, in fact, lead to progressive changes; but for reasons to be given later I consider that this but seldom occurs, that in fact the intercrossing of extreme forms must almost of necessity result in what might be termed retrogressive changes - result in something more than mere arrest in development. The crossing of two distinct varieties or of two well-marked species might very well be expected to result in an intermediate form; but owing to the antagonism between the units of germ-plasm derived from two dissimilar individuals, it is undoubtedly to many all but inconceivable that crossbred progeny should possess in an exaggerated form the characters of either of the immediate ancestors. We might a priori as well expect a mixture of red and blue sand to be intensely red or intensely blue rather than purple.
Nevertheless, the numerous stripes across the face and body of Romulus and the spots over the hind quarters may after all be due to discontinuous variation. Hence although Darwin was wont to ascribe to reversion the appearance of transitory stripes on foals, and the more permanent stripes on mules, it will be well to consider whether the zebra hybrids do not in their colouration afford a striking instance of discontinuous variation. Bateson in his suggestive work on variation says, "Around the term reversion a singular set of false ideas have gathered themselves," and that "it would probably help the science of biology if the word 'reversion' and the ideas which it denotes were wholly dropped, at all events until variation has been studied much more fully than it has yet been." ['Materials for the Study of Variation,' 1894.] He also says that to adopt the reversion hypothesis to explain the discontinuous occurrence of new forms possessing much perfection and completeness is inadmissible, because of the frequent occurrence by discontinuous variation of forms which, though equally perfect, cannot all be ancestral.
There is probably much truth in what Bateson says. Reversion may have been credited with results which could be sufficiently accounted for by variation. I have no means of knowing whether Bateson and others who have adversely criticised the reversion hypothesis, would account for zebra hybrids differing from their striped parents by discontinuous variation or not. As, however, reversion towards a remote ancestor in the case of the Equidae at least has not been proved - is probably not even susceptible of proof, - it is most desirable that we should endeavour to learn as much as possible from zebra hybrids for and against the reversion doctrine.
Theoretically, as already mentioned, the result of crossing distinct varieties or species should be the production of an intermediate form. This ideal result is seldom attained. It seems, however, that, as Liebscher has shown, the ideal result is sometimes practically reached, e.g. when the two species of barley, Hordeum stendellii female and H. trifurcatum male , are crossed, a hybrid is obtained which is as nearly as possible intermediate between the two parent forms. [Liebscber's work is referred to in Weismann's 'Germ-plasm,' p. 301.]
But usually, as Darwin long ago pointed out, the cross-bred offspring, instead of combining in equal proportions the characters of the sire and dam, may radically differ from the immediate ancestors and often closely resemble known or supposed remote ancestors. By inbreeding, the carriers of heredity or, let us say, the germ units responsible for maintaining the distinctive characters of any given race are fixed and it may be strengthened. Hence, when two inbred and closely related forms are mated, the offspring are likely to closely resemble their immediate parents. In cross-breeding exactly the opposite holds true; it is quite uncertain what the offspring will be like, unless one of the parents is highly prepotent. For example, if two closely related thoroughbreds are mated the offspring will almost certainly resemble the parents or the grandparents - the resemblance being greatest to the most inbred parent. But if a thoroughbred is mated with a cross-bred mare - with, say, a country-bred Indian mare - it is impossible to say what the progeny will be like; it may be large but quite devoid of quality, or small and fine, have plenty of quality but no substance. By inbreeding and by a form of selective breeding (which though quite a different thing may lead to similar results), new strains may be rapidly produced. Certain well-fixed types of Shetland ponies have been formed by close inbreeding, and similar results might be obtained without the disadvantages which follow inbreeding by judicious selective breeding. For example, if an English thoroughbred mare is mated with a thoroughbred horse whose ancestors for some generations have lived a comparatively natural life - in say Australia or America, - a horse, nevertheless, closely resembling the mare both in his physical and mental characters, the offspring would in all probability present all the striking peculiarities of the parents without any weakening of the constitution such as is apt to follow in and in breeding. But as allowance must always be made for variation (especially when forms from widely separated areas are mated), for the appearance of sports in one direction or another, the offspring might very well be an improvement on the parents- give evidence, in fact, of progressive development in some useful or desired direction, We often use the phrase "like produces like," but we often fail to realise that to give the offspring a chance of being like the parents, the parents or their grandparents must be like each other. It seems to me that, as a rule, progress in any given direction, i.e. progressive development, will only be possible when the parents closely agree with each other in their physical and mental peculiarities. They are, of course, likely to agree when closely related by birth, if their ancestors have been interbred for several generations; but they may also agree in all essential points when distantly related, or when belonging to quite distinct strains.
The opposite of mating closely related forms is the crossing of distinct breeds. The result of intercrossing is well known. We are all familiar with the phrase "the swamping effects of intercrossing," which implies that quite the opposite result is obtained from what we arrive at by isolation or inbreeding. Intercrossing, in fact, generally tends to arrest development in any given direction, to prevent the formation of new types or varieties; and hence, unlike inbreeding, it may be considered retrogressive in its tendency, and at the same time uncertain in its results. If the intercrossing of varieties in the same area arrests the formation of new species, is non-progressive, if not actually retrogressive, the crossing of distinct species from, it may be, widely separated areas will, a priori, not only arrest progressive development - abruptly put an end to the progress the respective species were making in various directions, - but actually lead to retrogressive changes, in other words, to reversion. The difference between breeding with almost identical inbred members of the same variety and with representatives (neither of them inbred) of two distinct varieties, may be made more evident by an example from amongst the pigeons. If two inbred fantails which present almost identical characters are mated, the offspring will in all probability agree closely with the parents, they may even show some advance in the direction desired by the fancier - in, say, the characteristic attitude or in the number of feathers in the tail. If, however, one of these fantails is afterwards mated with, say, a blue pouter, the young instead of being intermediate between a fantail and a pouter, may closely resemble the wild rock pigeon - may, in fact, have reverted towards the remote ancestor. This would simply be an exaggerated example of the swamping effects of intercrossing. If, therefore, the result of crossing extreme forms is, under ordinary circumstances, accompanied with retrogressive changes, I fail to see how the difference in the markings of Romulus from those of his sire could be considered as due to abrupt or discontinuous variation.
If in Romulus we have a new creation - a decidedly new variety rather than an attempt at the restoration of a lost ancestor - it is remarkable that he should so forcibly suggest in many ways the Somali zebra, and still more remarkable that he should in the plan of his markings not only agree with this year's crop of hybrids, but also with hybrids between zebra mares and various breeds of horses and to a considerable extent with zebra-ass hybrids.
If the result of crossing a blue pouter with four or five different recognised varieties of pigeons invariably resulted in the production of young resembling the blue rock, we should unhesitatingly account for this by the reversion hypothesis. If, on the other hand, the crossing led to new and quite distinct types that bred true, we should in time look upon these cross-bred forms as new varieties. In the same way, if the result of crossing zebras with four or five recognised species or varieties of the Equidae or horse family leads to the production of hybrids all marked on the same plan, we must, I think, admit that the resemblance is due in each case to reversion towards a remote - it may be a common - ancestor.
But if it is difficult to imagine how by discontinuous variation Romulus (with his numerous complete and perfect stripes on the face, neck, and body, and rudiments of stripes over the hind quarters) could be derived from a Burchell's zebra or an unstriped black mare, it will be found still more difficult to understand some of the markings in this year's hybrids. The new hybrids all differ from each other and from Romulus. Yet in no respect can they be said to present characters new to the Equidae; there is no evidence in any instance of new departures or of progressive development. The oldest (Remus), having a three parts bred bay Irish mare for his dam, though much lighter than the yearling, is marked in exactly the same way, but he possesses callosities (chestnuts) on the hind as well as the fore-legs, and his mane is as short and upright as in his zebra sire. About the next oldest (Heckla) there is nothing remarkable save that the body colour is already very dark. The dam is a skewbald (yellow and white) Iceland pony with white mane and tail. I expected her hybrid foal to be almost as light as a zebra, but instead the stripes are indistinct and the spaces between them mostly of a dark brown colour, while the mane and tail are nearly black. The hybrid of a small black Shetland pony - in some respects a small edition of Mulatto - is especially interesting. This hybrid filly suggests Romulus when at a corresponding age, but she is smaller and more compact and more self-assertive, but less docile. The general plan of the striping and the colouration is the same as in Romulus; and, as in Romulus, there are fourteen frontal arches (but the pattern is somewhat different), and there are nine vertical stripes across the body behind the shoulder stripe. Over the hind quarters, instead of numerous more or less transversely arranged spots, there have been from the first numerous narrow somewhat zigzag bands. In other words, unlike Romulus and the three other hybrids, in Norette the croup and rump have been from birth decorated by narrow transverse stripes such as we find on the Somali zebra. But although there have been since birth numerous distinct transverse narrow stripes extending well down the hind quarters in the Shetland hybrid, this is no argument in favour of discontinuous variation. In this case what might be termed the Somali stage in the ancestral history has been reached about a year sooner than was the case with Romulus.
The fact that all the hybrids differ so decidedly in the markings over the hind quarters from their common sire while they agree more or less accurately with the Somali zebra, seems to me to support the reversion hypothesis. Further evidence in favour of reversion is afforded by the neck stripes. Not very long ago, from a geological point of view, there were two types of three-toed horses in existence, one (Protohippus) in the New World, the other (Hipparion) in both the New and the Old. Protohippus was probably the ancestor of all our recent breeds of horses; Hipparion I look upon as the common ancestor of the asses and zebras. The asses I believe parted company at an early stage with the zebras - before the zebras had acquired their extravagantly striped coat. If asses were never as fully and distinctly striped as horses, hybrids between a common ass and a zebra might very well in their markings present a simpler and more primitive arrangement than hybrids between zebras and horses, more especially at points where in horses the stripes long persisted. Judging from the markings of my zebra hybrids, it might have been predicted that crosses between an ass and a zebra would present numerous irregularly arranged spots over the hind quarters, and numerous ill-defined wavy lines on the neck and on the sides of the body.
As a matter of fact, there is preserved in the Natural History Museum, South Kensington, a hybrid said to be between a common zebra and a common ass, which is profusely spotted over the hind quarters, and marked by ill- defined stripes on the neck and body. This hybrid has, as might have been expected, a distinct dorsal band, distinct shoulder stripes, and well-defined transverse bars across the legs.
If in these hybrids we have a more primitive form of marking than is ordinarily found in zebra-horse hybrids, it is conceivable that in some of my hybrids a more primitive arrangement of the stripes might be met with than occurs in Romulus; that as the mares belong to different breeds and colours, and differ in the extent of their inbreeding, the reversion (if there is such a thing) is likely to be more marked in some of the offspring than in others. I have already pointed out that though there are only twelve cervical stripes in Matopo (the sire of the hybrids) , there are, when the shadow stripes are included, over twenty in a Transvaal filly and twenty-four in Romulus. In the British Museum zebra-ass hybrid there are numerous stripes which, because of their branching and blending with each other, cannot be easily numbered. In the Shetland hybrid there is an intermediate condition of the neck stripes. Twenty-four stripes, as in Romulus, have been differentiated, but a number of indistinct stripes - like the shadow stripes in the zebra filly - occupy the intermediate spaces. That these narrow obscure stripes are the vestiges of never very distinct ancestral stripes, and that they correspond to some of the wavy lines in the zebra-ass hybrid, seems to me extremely probable. But similar "shadow" stripes also occur in a small bay Clydesdale mare's hybrid; only in this case obscure stripes are found between the vertical body stripes as well as on the neck. It is almost inconceivable that by variation there could be produced in a single generation not only what seems to be the typical number of neck and body stripes for a zebra-horse hybrid, but also shadow stripes, i.e. markings which mimic stripes in process of disappearing in some of the living zebras. By the reversion hypotheses their presence is easily understood, but to account for them by saying they are the result of abrupt or discontinuous variation seems to me to be alike unnecessary and illogical. Taking into consideration all the facts above enumerated, I am strongly inclined to the view that in Romulus and this year's crop of hybrids we have not new and striking variations, but more or less accurate restorations of certain remote ancestors, or an attempt to create the hypothetical "mid-parent" of the remote ancestry; in other words, we have strong evidence that under certain circumstances quite as pronounced reversion occurs in the Equidae as in pigeons.
It thus appears that when the colour and markings of Norwegian and other ponies are considered, the conclusion seems warranted that all the different breeds of horses have descended from yellowish-brown hog-maned ancestors, striped somewhat after the fashion of the mountain zebra (Equus zebra). Further, by taking into consideration the plan of the striping in the various living zebras, it may be inferred that of all the recent varieties and species the Somali zebra is in many respects the most primitive in the pattern of its decollation. It has also been shown that the five zebra hybrids already bred neither resemble their common sire, nor agree in their markings with Norwegian and other ponies, nor yet stand midway in colour, marking, &c., between their immediate ancestors. When due allowance is made for inbreeding in the respective dams, the conclusion seems inevitable that the remarkable differences between the hybrids and their immediate ancestors are due to reversion, either towards the less remote ancestors of the zebras or horses, or towards the common ancestors of all the recent Equidae. [Or to use Mr. Galton's phraseology, the hybrids have reverted so as to resemble the hypothetical "midparent," uniting the characters of their respective remote ancestors.]
In considering the question, Were the ancestors of the horse striped? it has been incidentally shown that all foals do not pass through a striped stage, and that many foals have stripes at birth which ordinarily completely or all but completely disappear. That foals are usually of a bay or dun colour, and frequently striped at birth, seems to be best accounted for by saying they wear for a time their ancestral colours, either owing to the influence of simple heredity, or to an extraordinary kind of heredity, commonly known as atavism or reversion. It has been especially shown that the greater the difference between the sire and the dam, the greater the reversion is likely to be; and, it may be added, the longer are the ancestral markings likely to be retained. For example, hybrids between two distinct species usually resemble the remote ancestors more than crosses between two varieties or races, and while cross-bred forms (crosses between closely allied races or varieties) may soon lose any hints they may have of remote ancestors at birth, hybrids generally retain the ancestral characters throughout life.
THE NATURE OF TELEGONY.
Having considered the question. Were the ancestors of the horse striped? and the still more important question, Does reversion occur in the Equidae? we are now in a position to deal more effectively with the telegony problem, as far as the horse family is concerned. It will be evident that by assuming horses have descended from striped ancestors, and that the less remote ancestors sometimes still reassert themselves, that by what we term atavism or reversion horses even now sometimes wear the ancestral colours, it will be evident that by granting these premises the question has been enormously complicated.
Lord Morton and his followers believed the Arab mare had been "infected," because her subsequent progeny to a black Arab horse had stripes and a more or less upright mane. If; however, purely-bred horses have sometimes a hog-mane and as many stripes as the Gore Ouseley "colts," how is it possible to prove that "infection" has occurred in any given instance? To take a concrete case, to what extent would Mulatto's second foal require to differ from an ordinary foal in order to prove "infection" has taken place? and, more especially, will the telegonous foal require to resemble Matopo (the previous sire) or one of Mulatto's or Matopo's less remote ancestors?
Hitherto experimenters seem to have invariably looked for some resemblance to the previous sire in the pure-bred subsequent offspring, and to have neglected or considered worthless resemblances to remote ancestors. The late Sir Everett Millais (who, as already mentioned, made all sorts of experiments during a breeding experience of nearly thirty years' standing, and made over fifty special experiments to induce a case of telegony for Mr. Romanes) records what he considered a perfectly authentic instance of telegony. ['Two Problems of Reproduction,' p. 20.] In this case a pure-bred fox terrier "was spotted exactly like a Dalmatian." The litter previous to the one in which the spotted fox terrier appeared was sired by a Dalmatian, and hence it was inferred that "infection" had taken place, the result being in this case a striking resemblance on the part of the subsequent off- spring to the previous sire.
At a meeting of the Zoological Society held on November 17th, 1896, "Mr. Chalmers Mitchell exhibited and made remarks on a supposed case of telegony as shown by a fox terrier whose mother, a pure fox terrier, had previously bred some mongrel dachshunds." Sir Everett Millais "denied that the specimen exhibited any trace of the influence of the dachshund sire of the previous litter, and traced the divergences of the puppy from the modern fox terrier type to the original source of the breed." [The Field, Nov. 28th, 1896.] In other words, Sir Everett was evidently not prepared to admit that "infection" might lead to the subsequent progeny resembling a remote ancestor - "the original source of the breed."
In support of my statement that the previous sire has been hitherto taken as the standard for comparison rather than a remote ancestor, the following cases may be cited:
(a) A black and white sow which had offspring to a chestnut-coloured wild boar had afterwards a litter to a boar of her own breed, some of which were marked with chestnut, ['Phil. Trans. Roy. Soc.,' 1821, p. 21.]
(b) A dachshund which had a litter to a sheep-dog had afterwards pups to a pure dachshund; the second and third years' produce resembled the sheep-dog as much as the dachshund sire. [Herbert Spencer, Contemp. Review, May, 1893.]
(c) An ordinary cat had kittens to a tailless Manx cat, and then to a cat of her own kind; several of the second litter of kittens were tailless. [Journ. des Debats, Sept. 9th, 1897.]
(d) "Blair Athol," a blaze-faced chestnut horse, according to various writers "infected" the mares that bred to him, with the result that some of these mares produced Blair Athol like foals to "Wild Oats"- "the last horse in the world to get stock of a similar stamp," i.e. to get Blair Athol like foals. [The Sportsman, Jan. 3rd and Feb. 29th, 1896, letters by the "Special Commissioner."]
If a telegonous foal resembled, as has generally been assumed, the previous sire, while an atavistic foal of necessity takes after a more or less remote ancestor, it would be comparatively easy in many cases to say whether infection had taken place, to decide on the lines adopted by Sir Everett Millais in the case of Mr. Chalmers Mitchell's fox terrier. If, on the other hand, the result of "infection" is identical as far as it goes with the result of reversion, coming to a definite conclusion will be difficult - will seem, in fact, to some next to impossible.
I had not intended to consider at this stage the possible causes of telegony, but as the result of "infection" (if it occurs) will depend more or less on the method by which it is produced - in other words, as the mode of "infection" may determine whether the subsequent offspring resemble the previous sire or a remote ancestor, it is desirable to refer briefly to some of the suggested explanations of the supposed phenomenon.
Sir Everard Home believed Lord Morton's mare produced striped foals to the black Arab horse because her first mate - the quagga - had produced a profound and lasting impression on her nervous system; that she, as it were, had. never quite succeeded in getting the quagga out of her mind. Sir Everard tells us he considered the markings on the subsequent offspring of Lord Morton's mare as "one of the strongest proofs of the effect of the mind of the mother upon her young that has ever been recorded." ['Home Lectures on Comparative Anatomy,' vol. iii, p. 307, 1823.] If telegony is due to mental impressions, as Sir Everard Home and many others since his day have believed, the subsequent progeny might very well resemble (I might almost say ought to resemble) the previous sire, and not a remote ancestor. But although it is widely and firmly believed that the offspring may be influenced through "the mind of the mother," Sir Everard Home's is the least likely of all the explanations hitherto suggested.
Mr. Herbert Spencer believes telegony is due to a kind of pangenesis, that "while the reproductive cells multiply and arrange themselves during the evolution [development] of the embryo, some of their germ-plasm passes into the . . . parental body," to eventually reach and be after- wards included in the reproductive cells subsequently formed. [Contemporary Review, March, 1893.] This explanation has found favour with not a few physiologists in the past, and is still seriously considered. But even granting the possibility of this indirect mode of reaching and infecting developing (maturing) reproductive cells, it by no means follows the offspring- resulting from these cells will resemble the previous sire. All that could be safely asserted is that the subsequent offspring would resemble the previous offspring, or the ancestors of the immediate parents. If, for example, Mulatto's germ-plasm was indirectly infected by germ- plasm from her first foal during its development, her second foal to the Arab horse (granting telegony is true) should more or less resemble her first foal (Romulus), or one of her remote ancestors, rather than her first mate Matopo.
Professor Weismann thinks telegony (if it occurs) is best accounted for by supposing that some of the germ-plasm of the first sire reaches and penetrates some of the immature ova as well as the mature one from which the first embryo is developed." [Weismann, 'The Germ-plasm.'] This germ-plasm, instead of merely serving as so much nourishment to these immature ova, according to this view retains its independence and eventually asserts its influence on the future progeny to a second and different sire. This may be better understood by an example. If any given simple unicellular organism were to devour another but smaller organism belonging to a different variety or species, and then unite with one of its own species, when division eventually took place the new individuals would (supposing the comparison to hold) present some of the characters of the species originally devoured as food. Although it is difficult to understand how an egg could incorporate germ-plasm in this way while still in pro- cess of maturing, and still more difficult to understand how this incorporated germ-plasm could retain for an indefinite time its independent existence (play the part, as it were, of an encysted cellular parasite, which at the right moment wakes up and insists on being represented in the future offspring), this explanation, regardless of the fact that vestiges of male germ-cells have never been seen in immature ova, will evidently be very generally adopted should telegony be eventually raised to the rank of a fact. [Mr. Heape evidently believes that if telegony occurs, the characteristics of the first sire can only be transmitted to the offspring of the second through the germ-plasm of the dam. - Nature, Dec. 30lh, 1897, p. 215.]
If telegony is due to the direct "infection" of the germ-plasm of the dam by the germ-plasm of the first or of a previous sire, there seems no escape from the conclusion that the subsequent progeny should resemble the first offspring rather than the previous sire. In the case of Mulatto, her second foal (to the Arab horse) should, if infected, resemble her first hybrid foal Romulus. I have already pointed out at considerable length that Romulus is surprisingly unlike his Burchell zebra sire, and I have given reasons for the belief that he takes after the Somali zebra, probably the most primitive of all the living striped horses. Hence, if there are stripes in Mulatto's second foal, it might be argued they should of necessity agree with the stripes in Romulus or in the Somali zebra or in some of Mulatto's ancestors.
In other words, the result of "Infection," according to the generally accepted (and it must be confessed most feasible) explanation, should as far as it goes be identical with "Reversion." It was in anticipation of this conclusion that I was led to discuss so fully striping and reversion in the Equidae, and to consider experiments with dogs at the out- set of comparatively little value seeing we know so little of the dog's ancestors. But if the developing ova are capable of incorporating rather than assimilating germ-plasm after the manner suggested by Weismann and Romanes, why is telegony so extremely rare? - according to Romanes only occurring in one or two per cent, of cases; according to Sir Everett Millais not only exceedingly rare but abnormal.
Three reasons occur to me why telegony, if possible, happens or rather appears to happen but rarely. In the first place, the germ-plasm of the previous sire may only be able to obtain a footing in ova at a certain stage of ripeness. If germ-plasm (of the first sire) enters eggs before a certain stage in the process of maturation is reached, it may serve as so much nourishment, and thus fail to assert itself in the subsequent progeny; whereas if it enters later it may obtain a footing, reach the nucleus, and eventually take part in the formation of the subsequent offspring. In the second place, the whole or almost the whole of the incorporated germ-plasm of the previous sire may sometimes be discharged in the polar bodies, i.e. when the nucleus of the all but ripe egg is discharging the half of its germ-plasm. Were this to happen the "infection" would either not take place at all or be so limited that it might readily escape notice. In the third place, telegony may seem to be rare because of the prepotency of the sub- sequent sire. The first two explanations are at the best mere guesses, but the third may be a very real explanation.
It is well known that some breeds are so fixed that they almost invariably produce offspring like themselves, however they happen to be mated. For example, while the black Galloway polled cattle almost invariably throw blue-greys to white shorthorn bulls, they frequently yield to other breeds offspring so like themselves that even experts are at times deceived. For example, "Twenty years ago the late Duke of Buccleuch, K.G., tried the experiment of mating Galloway bulls with West Highland heifers. The bulls he used were Baron Douglas (614) and Border Chief of Drumlanrig (1015). The females were superior West Highland heifers, one of which, I understand, was bred by Mr. Stewart, of Ensay, and the other by Lord Malcolm, of Poltalloch. When the produce - two heifers - were grazing, at the age of about eighteen months, among a lot of nearly a score of pure-bred pedigree Galloway heifers, half a dozen of the most experienced and best known breeders of Galloways were asked by his grace's manager to pick out the Galloway-Highland crosses from among the pure ones, and each of these experienced judges picked out the wrong animals, so closely did the half-breeds in every particular resemble the pure ones." [From a letter by the Rev. John Gillespie, LL.D., president of the Highland and Agricultural Society, the Field, January 1st, 1898.] Mulatto's hybrid Romulus might hence have been the image of his gaudily- painted sire Matopo. It is even conceivable Romulus might have closely resembled his almost jet-black dam.
When hybrids or cross-bred offspring are identical or nearly identical with the first sire, the "infected" subsequent offspring (telegony being taken for granted) might also resemble the first sire. I say might advisedly, because if the second sire happened to be distinctly inbred, or for some other reason extremely prepotent, the influence of the dam, plus the influence of the first sire, might be completely overcome, with the result that the subsequent progeny would very closely resemble the second sire.
It thus appears that the further the telegony problem is considered the more complicated it becomes. But not only has inbreeding to be taken into consideration, the fact already insisted on must also be borne in mind, viz. that pure-bred foals are occasionally striped; in other words, that the result of "infection" on the subsequent progeny by a zebra sire may be identical or almost identical with simple reversion.
If infection is not only possible but constant, it follows from what has already been said that the telegonous offspring might either (1) resemble a remote ancestor, or (2) the previous sire, or (3) the subsequent sire, or (4) the dam.
[* It should be borne in mind that any part of an animal may be "infected;" that in the case of foals evidence of infection may be found in, e.g., the hoofs, though not in the warts;' in the structure, though not in the colour of the hair; in the movements and habits, though not, as far as the eye can detect, in the muscles or nervous system. Hitherto attention has been mainly directed to the colour of the hair and feathers in experiments bearing on reversion - hardly at all to the general structure and habits. That crossing of somewhat extreme forms affects the nervous system, the muscles, and skeleton, as well as the colour and form of the skin and its appendages, hair, feathers, hoofs, &c., is extremely probable. Last summer a small dark bantam hatched out nine chicks, three of which were by a large Indian game cock having some Dorking blood in his veins. Of the three cross-bred chicks the two survivors might now be almost taken for jungle fowl, the cock only differing from some pure- bred birds I saw recently at Achnamara, in Argyllshire, in having a double comb and comparatively few dark hackles on the breast. But what is perhaps more remarkable, the birds resembling jungle fowl in size and colour differ from the others in their habits. For example, when suddenly disturbed they fly off, sometimes for a considerable distance, evidently in a state of great "alarm; while the other members of the same brood either take little notice of the intruder, or simply run under a hedge or whatever cover may be available. McKenny Hughes, in his memoir [‘Archselogia,' vol. Iv, pp. 125-158.] on "British Breeds of Cattle," refers to numerous horn-cores found in ancient ditches (twelfth to fifteenth century) cleared out for the foundations of the University Press and other buildings in Cambridge during the years 1892-3. Of over 125 horn-cores "there were none that could be referred to a long-horned breed, and few that showed any traces of Roman type." After carefully considering the matter, McKenny Hughes came to the conclusion that these horn-cores afforded evidence of the existence of a mixed breed in the act of "reverting sporadically to various primeval types, but principally to the numerically predominating Bos longifrons," i.e. to the small ancient British (Celtic) shorthorns.]

Fig. 40. - Mulatto.
Fig. 41. - Benazrek.
From the fact that Romulus resembles the Somali zebra, Mulatto's second foal should also, though only to a limited extent, resemble the Somali zebra, - or, which is the same thing, somewhat resemble Romulus. Had Matopo been inbred and more prepotent than Mulatto, then Romulus would in all probability have resembled Matopo. If, conversely, Mulatto had been inbred and more prepotent than Matopo, Romulus would most likely have resembled Mulatto. Mulatto's second foal, if to a non-prepotent sire, should resemble Romulus; but if to an inbred prepotent sire, the second foal ought rather to resemble the second sire. If, however, the second sire belongs to a different breed from Mulatto, - if there is a certain amount of antagonism between the germ-plasms, - the second foal may revert, and resemble one of the less remote ancestors of the true horses.
[* Many who believe telegony occurs in mammals seem to think it is impossible in birds. If telegony is due to indirect "infection," to germ- plasm proceeding from the developing embryo by a roundabout road to ova in process of maturing, one can understand why in birds it is impossible. If, however, the germ-plasm of the first sire passes directly to the immature eggs, there is no reason why there should not be telegonous birds as well as telegonous mammals. Sir Everett Millais experimented with ducks, hens, and pigeons, without obtaining any evidence of telegony. If, however, he only looked for a resemblance to the previous sire, or if the subsequent sires were highly inbred, cases of infection may have been overlooked or obscured. For example, if a white fantail hen is first bred with a blue pouter and next with a white fantail, one of two things might happen, - the offspring to the second mate (the white fantail) might be identical with their parents (if they happened to be inbred), or suggestive of a blue rock pigeon. If the latter result happened, it would be considered by some a case of reversion, regardless of the fact that the first cross between a white fantail and a blue pouter might (through the influence of reversion) resemble not a pouter but a rock pigeon.]
It would obviously be impossible or all but impossible to distinguish this form of reversion - reversion resulting from crossing two distinct breeds - from reversion due to infection by a previous sire. There is still another possible, it may be a highly probable, result. The influence of the previous sire may be so feeble that, instead of leading to a reversion towards the hypothetical "mid-parent" of the remote ancestry - that is to offspring combining to a limited extent the characters of the ancestors of both the horses and the zebras, - it may only set up sufficient disturbance in the reproductive system of the dam to lead to a slight regression towards her own particular and not very remote ancestors.
Before describing Mulatto's second foal, the question may be asked, what amount of striping or other changes would be required to conclusively prove she has been "infected" by the zebra? Obviously if the germ-plasm of the first sire unites with a maturing egg, the previous sire could only enter into the formation of the second foal to a very limited extent. According to Mr. Galton's law of ancestral heredity, the two parents contribute one half, the four grandparents one fourth, the eight great-grand- parents one eighth, and so on, to the total heritage of the average offspring. If this law holds it follows that the male parent only contributes one fourth of the total heritage. If the previous sire contributed equally with the reputed sire, the proportion would be one eighth for each. But even if we admit that an immature egg may contain the whole of the male germ-cell of a previous sire, i.e. one eighth, this would be halved during the "reducing division" of the nucleus; hence, at the most, the contribution of the previous sire would be one sixteenth, but actually (supposing there is direct "infection" of the ovum) in the vast majority of cases considerably less. Further, the half not contributed by the parents would be mainly provided by the ancestors of the dam. Hence, when the most liberal allowance is made, the previous sire could only under ordinary circumstances enter to a very limited extent into the subsequent offspring. It is true that the colt and filly bred by Sir Gore Ouseley from Lord Morton's mare were more richly striped than the quagga hybrid. This has always seemed to me to prove that the majority of the stripes in the Gore Ouseley "colts" were inherited directly from the black Arabian.
Mr. Wilfrid Blunt believes that the offspring of white Arabs are apt to show stripes; while Herr W. von Nathusius, in a recent letter, indicates that he thinks there are latent stripes in black horses. From what I have seen of black horses when in the act of shedding their winter coat, I think one may go further than Nathusius and say that stripes sometimes actually exist in black or all but black horses. Supposing there is no such thing as telegony, and that white and black horses tend to produce striped offspring, and, further, that when fairly distinct types are crossed the offspring are likely to revert, what extent of striping might be expected in the foal of a black Island of Rum pony which has never bred with a zebra, and a grey Arab horse?
Darwin tells us he "never heard of either shoulder or leg stripes without the spinal stripe. The latter is by far the commonest, as might have been expected as it characterises the other seven or eight species of the genus." ['Animals and Plants,' vol. i, p. 68.] This has also been my experience, and, as far as I know, it has been the experience of all who have directed their attention to the subject. Hence Mulatto's second foal (even if there is no such thing as telegony) might have a dorsal band, shoulder and leg stripes, and a number of not very distinct stripes on the face, neck, and body. We still know too little of the plan of the stripes in the ancestors of the horse to admit of their being compared with the stripes in the zebras; what has been ascertained, however, points to the arrangement differing considerably. The shoulder and leg stripes so common iu the domestic ass may be said to be transmitted almost unaltered to zebra-ass hybrids. There is, however, no evidence that any stripes are derived directly from the horse in zebra- horse hybrids; it looks as if the stripes of the ancestors of Mulatto had been overcome or masked by stripes characteristic of the ancestors of the zebras.
The likelihood of stripes in addition to shoulder and leg stripes and a dorsal band occurring iu a foal bred from an Island of Rum pony and a grey Arab is extremely small. The probability, e.g., of croup stripes appearing- is not great, for though in some foals the hair across the croup forms wavy bands, stripes in this position are extremely rare. I am not forgetting that Mr. Darwin bred a foal marked with numerous narrow stripes across the croup; but in the absence of a figure it is impossible to say whether these narrow stripes were comparable to the stripes found in say the Somali zebra, or in some of my zebra hybrids. Hence, as already said, the probability of an Island of Rum pony which had not been crossed with a zebra producing a colt striped over the croup is not very great.
MULATTO'S SECOND FOAL.
The second foal of Mulatto was born on the 16th July, 1897, the period of gestation being 330 days - twelve days shorter than in the case of Romulus. The dam (Mulatto, Fig. 40) has already been sufficiently described. The sire (Benazrek, Fig. 41j, bred by the late Earl of Warwick, is a grey Arab, a son of Azrek, for some time a member of Mr. Wilfrid Scawen Blunt's famous stud at Crabbet Park, Sussex. Azrek, also a grey, belonged to the ancient Seglawi Jedran family. Benazrek's dam (Shemptes) was also originally a grey; now she is almost white. I have had under observation for some time three "colts" by Benazrek. One, having a well-bred bay mare for her dam, was of a reddish dun colour at birth, with a blaze. This "colt" (now - 1898 - rising three) will ultimately be grey like her sire. Even at birth there were no indications of stripes. A second foal by Benazrek (also rising three) had for her dam a dark grey thoroughbred Irish pony. This foal was at birth of a dull chestnut colour, but now she is a dark grey not unlike her dam. Like the above- mentioned foal, this filly has never shown any stripes. Benazrek's third foal (born May 30th, 1897) has for her dam a flea-bitten 13 hands New Forest pony, in which there may very well be a trace of Arab blood.
This pony had a mule foal (Fig. 42) to a Forest donkey in 1895, but missed having a foal in 1896. At birth her foal to Benazrek was of a reddish dun colour; when three months old the foal was of a light grey colour; at eight months a dark grey, with ablaze and a small reddish patch near the left nostril. This foal when a few days old had a number of stripes. There was a distinct dorsal band, a stripe nearly two inches in width across the withers in the position of the shoulder stripe in the common ass, and in front of this on each side of the neck two narrow cervical stripes. There were also three faint vertical stripes extending downwards from behind the withers to a short obscure ventral band; further, there were obscure bars above the knee and hock, and across the chest and the upper part of the forearm. When a month old the leg stripes were no longer visible; when two months old the neck stripes had vanished and only a vestige of the shoulder stripe was left; but even at three months the vertical stripes extending down behind the elbow were quite evident.

Fig. 42. - Mule. Dam a flea-bitten New Forest pony.
Only an indistinct dorsal band now remains. Whether these stripes are due to the previous sire being a donkey it is difficult to decide. In favour of the view that the Forest pony has been infected by the previous but by no means first mate are two facts worth mentioning. I have already given reasons for the belief that in the remote ancestors of the horse long hairs were confined to the lower part of the tail, and that it is only during comparatively recent times that long hairs have extended up to near the root. It is known to many that in most horses the hairs from the upper two or three inches of the tail are shed with the winter coat. But the shedding in the vast majority of cases is so gradual, and the growth of the new hair so rapid, that the loss of the dead hairs is hardly perceptible. In the mule, on the other hand, the long hairs from about six inches of the root of the tail are usually shed so rapidly that only a covering of comparatively short hair is left. My zebrules agree with ordinary mules in this respect, but the shedding of the long hairs may be more extensive, only the long hairs springing from the lower third of the dock persisting. In Hyla, the Forest pony's 1897 foal, all the long hairs of the upper 2-and-a-half inches of the tail were shed during April and May (1898), leaving the base of the dock almost bare. In the sire and dam of Hyla the hairs are shed very gradually from the root of the tail - a casual observer would never notice that the old hairs are being slowly replaced by new. The other fact is that the wart (chestnut) is absent from the left hind leg. Warts have never, as far as I am aware, been found on the hind legs of asses or zebras, and they are generally believed to be invariably absent from the hind legs of mules. But they are also said to be sometimes absent from the hind legs of horses. In the Forest mule (Fig. 42, Hyla's half-sister) warts, as it happens, are present; or, to be more accurate, there are small, smooth, bare spots in the position of warts. In all probability many mules have vestiges of warts, and it is just possible that in horses said to be without warts there may be similar vestiges. In one of my zebrules there is a small wart on one of the hind legs. The absence in Hyla of a wart from one of the hind legs, the falling out of the hair from the root of the tail, and the presence of faint indications of stripes across the croup, may be looked upon as mere accidental coincidences, or as the result of simple reversion; but, on the other hand, these unusual conditions may be due to "infection" - to, let us say, some of the germ-plasm of the previous sire (the Forest donkey) having asserted its influence more particularly in the tail, warts, and hind quarters.

Fig. 43. - Mulatto's Second Foal.
Fig. 44. - Mulatto's Second Foal.
Mulatto's second foal, when seen in bright sunshine half an hour after birth, seemed to be quite devoid of stripes. When, however, the foal was carefully examined in a suitable light, numerous stripes revealed themselves. The foal was of a dark bay, in some parts almost of a brown colour, and as the majority of the stripes were only a shade darker than the rest of the coat, they were, except in a favourable light, difficult to see and almost impossible to photograph. When a week old Mr. Edwin Alexander" was kind enough to fill in the more important stripes in an outline sketch, a photograph of which is reproduced in Fig. 43. [To Mr. Alexander, jun. (already noted for his accurate and beautiful drawings of animals), I am greatly indebted for the sketch (Fig. 43) of Mulatto's second foal.] I carefully compared the sketch with the foal, and had no difficulty in making out all the stripes shown in the figure. While some of the stripes were quite distinct to the eyes of even unskilled observers, others were most subtle and only visible in certain lights. Fig. 44 has been reproduced from a photograph taken when the foal was ten days old.
In the zebra the hairs of the dark stripes and bands are pigmented from root to tip. In Romulus the dark hairs are as a rule only pigmented to within about half an inch of their roots, whereas in Mulatto's second foal there was only a faint change of colour near the apex of the hairs forming the indistinct stripes. If the stripes in this foal are due to "infection," the hairs forming them could hardly be expected to be as deeply and extensively pigmented as in the half-zebra Romulus. As Fig. 43 shows, there are cervical but do shoulder stripes, numerous stripes on both the fore and hind legs, vertical stripes across the body, oblique stripes across the loins, faint stripes across the flank feather, and, most interesting and important of all, transverse stripes across the croup. I have already mentioned that Darwin never found leg stripes occurring in horses without the dorsal stripe, and even in the striped colt he bred there was a trace of the spinal stripe. In Mulatto's second foal, however, no trace of a dorsal stripe could ever be detected, i.e. one of the oldest if not quite the oldest stripe in the Equidae family was not represented in this foal. The stripes on the neck and the lower parts of the legs are not specially significant, but the stripes between the elbow and the shoulder-joints and on a level with the stifle are undoubtedly rare even in foals, while numerous narrow stripes in the region of the loins and stripes across the croup have, as far as I am aware, only previously been observed in the foal bred by Darwin.
In Darwin's foal, it will be remembered, "almost the whole body was marked with very narrow dark stripes, in most parts so obscure as to be visible only in certain lights, like the stripes which may be seen on black kittens." These stripes were plainest on the forehead, where they formed a set of pointed arches one under the other, but they were also "distinct on the hind quarters, where they diverged from the spine and pointed a little forwards." This colt ultimately turned out brown, but when "two or three months old all the stripes entirely disappeared." ['Animals and Plants,' vol. i, 2nd edit., p. 60.]
In the absence of a figure it is, unfortunately, impossible to contrast the stripes described by Darwin with those in Mulatto's second foal. I may, however, point out that narrow stripes or, to be more accurate, narrow ridges (some of them over, some under, a quarter of an inch in width) occur in all foals, more especially over the loins and croup and in the vicinity of the flank feather. These ridges are especially evident at birth in well-bred foals having a fine coat.
When the finger is carried across these ridges, one is tempted to conclude they are entirely or mainly due to crumpling or wrinkling of the skin, I was led to suspect they were not due to crumpling of the skin by noticing how the hair was shed in an Irish bay mare (Biddy) and in her yearling hybrid, Remus, In the latter, the spaces between the stripes across the hind quarters first lost their hair, with the result that the stripes projected freely for some time above the surface. In the dam, the hair in the vicinity of the flank feather was shed so as to leave a number of narrow tracks having the same disposition as the ridges in new-born foals. Eventually the hair forming the tracks was shed, leaving a perfectly smooth coat. Soon after noticing this I had sent me a well-bred foal (which died when about a week old) showing the characteristic ridges on the sides of the body as well as over the croup, loins, and hind quarters. I succeeded in getting the skin preserved without in any way altering the appearance of the ridges in question. When this skin was held up to the lights it was at once evident that the ridges were not due to furrows in the skin, but to the presence of tracks of long hair, separated from each other by narrow spaces sparsely covered by short fine hair.
About the same time I had the opportunity of examining twins that had been born nearly two months before their time. [Sire of twins a Clydesdale: dam a light cart-horse.] In one (the smaller) there were over the trunk narrow tracks of long and relatively thick hairs, having the same general direction as the hair in the adult. The spaces between these tracks - on an average half an inch in width - were occupied by short, fine, and almost invisible hairs. In the larger twin, each track was wider but less evident, owing to the intermediate spaces being better clothed. The ridges in the young foal and the rows of relatively long thick hairs in the foetuses agree very closely, if not absolutely, with the lines of moisture seen on a horse when it begins to perspire, and with the lines followed by the water after a light shower. In the prematurely born foals, two phases in the evolution of the coat of the horse may be represented; the least developed seems to point to a time when there was but a sparse covering of hair, but yet with the hair so arranged that it served to carry off the rain as rapidly as possible along definite lines.
When the skin showing the narrow parallel ridges and furrows is examined, the mane is seen to extend nearly to a level with the flank feather. From the withers the hair of the mane gets gradually shorter, but it runs in the same direction and has the same structure as the hair forming the mane in front of the withers. From the part of the mane behind the withers, ridges quite a quarter of an inch wide run downwards and slightly backwards over the sides. Over the croup there is a smooth patch of hair of a lozenge shape, measuring about six inches by one and a half at the widest part - a patch corresponding to the wide portion of the dorsal band in zebras and horses. From the front part of the lozenge about fourteen narrow ridges arch outwards and downwards in front of the flank feather (Fig. 45); while from the back part equally narrow ridges arch backwards over the hind quarters - the upper ones approaching each other as they proceed towards the root of the tail. Filling up the space between these two sets of ridges are others which spring from the flank feather, some bending downwards in front, others behind, the feather, towards the stifle. Fig. 45 indicates the arrangement of the ridges around the upper end of the right flank feather. Fig. 46 shows the appearance the hair presents in the space immediately above the flank feather (lying within the four circles shown in fig. 45) when the skin is held up against the light. In a week-old cinnamon-coloured foal by a thoroughbred chestnut horse out of a bay half-Arab mare, the flanks, croup, hind quarters, and thigh are marked all over by very distinct "narrow stripes," some of which appear of a different colour from the intermediate spaces. But all these apparent stripes, whether they run parallel with or at nearly right angles to the direction of the hair, are due to almost hairless spaces separating relatively long and thick ridges.
45. Ridges formed by hair in region of flank feather in a foal
46. Part of skin shown in Fig. 45
In addition to forming narrow ridges and furrows, the hair may be so disposed in some foals as to produce wavy bands across the croup, and faint lines at nearly right angles to these across the hind quarters. The fairly wide bands across the croup I have especially noticed in half-bred bay foals. These bands, which are never due to pigment, and are only seen in certain lights, fairly accurately correspond in their position with the cross-bars of the gridiron of the mountain zebra.
Evidently, in considering whether any given foal is striped, the narrow tracks of hair, especially evident in the vicinity of the flank feather, the wavy bands across the croup, and the faint bands across the hind quarters, will have to be allowed for. Coming to Mulatto's second foal, it may first of all be mentioned that there were no stripes on the face or across the shoulders, nor yet was there a dorsal band; and though there were bars across the legs, the bars were not so pronounced as in many purely-bred foals - foals whose parents had never seen a zebra. More- over, the leg bars were least distinct where they are generally most marked in dun-coloured ponies, i.e. in the vicinity of the knee and hock. There was thus an all but complete absence of the stripes that are ordinarily found in the horse. In Lord Morton's "colts" besides a dorsal band, there were "dark stripes across the forehead and dark bars across the back part of the legs," and stripes across the withers; in the filly the stripes covered "nearly the whole of the neck and the back as far as the flanks." There were, in fact, more stripes on the filly than on the quagga hybrid. It seems to me that at least all the stripes found in the filly over and above those in the quagga hybrid must be credited to the black Arabian, and not to the previous sire - the quagga. Why in Mulatto's second foal the stripes were not developed along the spinal ridge and across the withers and hocks, and were specially distinct across the croup and the front of the arm, I am unable to say. That they failed to appear in the usual position, and were more or less distinct in unusual positions, may be considered by some as evidence in favour of "infection." The stripes on the neck and legs need not be specially referred to, but the stripes across the croup and hind quarters deserve very special consideration.
In zebra-ass hybrids there are often numerous spots over the croup and hind quarters. These spots were present in a zebra-ass hybrid bred at Windsor during the reign of George IV, and also in a hybrid between an Asiatic ass and a zebra mare bred some years ago in Paris. In Romulus there were, at birth, spots over the croup, but in course of time the spots united to form somewhat zigzag stripes. In one of the 1897 hybrids there were narrow bands over the ci'oup from the first. In Matopo the croup and hind quarters are almost entirely occupied with the three flank stripes.
Evidently the bars across the croup in Mulatto's second foal have not been inherited directly from the first sire - Matopo. Their presence might be accounted for by saying they were latent in Mulatto, and that mating her with a horse of a different strain led to their development. But the offspring of the bay mare and of the dark-grey pony by the same Arab horse, showed no markings across the hind quarters. Is this because bays and dark greys are less liable to produce striped offspring? If the croup stripes in the second foal were not inherited from or induced by the Arab horse, they, it may be argued, can only be explained by saying that Mulatto has been in some way influenced by her first mate - the zebra Matopo.
It thus appears that in Mulatto's second foal, (1) the stripes which most frequently occur in horses were absent; (2) that there were certain stripes which are not uncommon in horses; and (3) that the most distinct markings present, viz. the stripes across the croup, are extremely rare in both foals and horses. While some of the stripes can easily be accounted for by simple reversion, it is difficult to account for others in this way.
In favour of the croup stripes having resulted from a simple reversion due to the crossing of somewhat diverse types, the experience of Darwin may be advanced. His striped colt was of a mixed origin. The dam was a descendant of a dark brown Flemish mare by a light grey Turcoman horse. The sire was Hercules, a thoroughbred dark bay, whose sire (Kingston) and dam were both bays. If the crossing of distinct types leads to reversion, we can well understand Darwin's striped colt being an example of reversion; and in the same way the striping in Mulatto's second foal might also be due to mating more or less distinct strains. Again, from the fact that the second foal had stripes across the croup instead of rows of spots as in Romulus, it might be argued that they could not be due to the influence of the previous sire. The difference in the marking of the croup between the first and second foal might, however, be explained satisfactorily by saying that in the one case the antagonism between the units of germ-plasm had been greater than in the other, with the result that the half-zebra Romulus reverted further than the second foal, in which the zebra could only be represented to the extent of about one part to thirty- two of the horse.
From various experiments made with dogs, fowls, &c., it is evident the amount of the reversion varies greatly, e.g. while some of the cross-bred fowls already referred to resembled not a little jungle fowls, others took after their immediate maternal ancestor, the dark bantam. Again, the offspring between a well-bred yellow collie and a Dalmatian were not whole-coloured or wolf-like in appearance, nor yet were they spotted like the sire. They were, in fact, extremely like foxhounds; on a white ground colour there are in each case several large brown blotches. In this case presumably the reversion has gone the length of the less remote ancestors of the Dalmatian (said to be pointers), but no further.
In favour of the croup stripes being due to "infection" or telegony is the fact that, with the exception of the New Forest pony's foal, Benazrek's offspring have hitherto been quite devoid of stripes; and the New Forest pony, having borne a mule foal to a jackass, may very well have been "infected." There is, however, a still stronger argument, viz., that the percentage of chances is against Mulatto producing a foal striped like Darwin's colt. I have been unable to find any records of foals characterised by stripes over the croup or rump in addition to Darwin's; and of the many experts who have examined Mulatto's second foal, not one had ever seen a colt so richly striped. It is, of course, possible that thousands of foals have been born with stripes over the hind quarters and loins - stripes so indistinct that they escaped notice. If pronounced reversion towards a striped ancestor is common, then there is nothing remarkable about the second foal; it is quite unnecessary to search for an out-of-the-way explanation, such as telegony supplies. If, on the other hand, croup stripes are extremely rare, if they only occur once in say every ten thousand foals, the chance of all the stripes on the second foal being due to simple reversion is extremely remote. Again, the Island of Rum ponies have been under observation for many generations, and yet there is no record of their foals ever having more than a dorsal band and a cloudy patch over the withers.
Further, it may be pointed out that it is not a little remarkable that both Lord Morton's and Lord Mostyn's mares, after having borne quagga hybrids, produced somewhat zebra-like foals to horses. Had Mulatto not been first mated with a zebra, she, like the grey Irish pony and the well-bred bay mare, might have produced a foal devoid of stripes, or with, at the most, a dorsal band and obscure bars on the legs.
I may mention the mane, hoofs, and tail of Mulatto's second foal were, as far as I could judge, perfectly horse-like; that the eyes were hazel-coloured, as in Matopo and in most of the West Highland ponies; while the eyelashes consisted partly of straight hairs, as in Mulatto, and partly of the characteristic-curved hairs as in the zebra hybrids.
Although as the hair increased in length and the foal's coat was shed the markings grew fainter, the croup stripes could still be detected at the end of December, when, unfortunately, Mulatto's second foal succumbed to ailments resulting from the presence of countless numbers of the parasite Strongylus. Although evidence of reversion could hardly be expected in the skeleton, I have examined the condition of the ulna and of the second and fourth digits. The ulna I found still complete, as complete as in an eight months' foetus; and with the exception of about an inch (near but not quite at the lower end) completely ossified, and affording a large articular surface for the cuneiform or outer carpal bone. Further, the inner vestigial digits, i.e. the vestiges of the second digits of both the fore and hind limbs, are relatively large, although there is no indication of either the middle or distal articulations. In concluding this contribution to a difficult and complex subject, I need neither point out that it would be premature to come to any conclusion as to whether there is such a thing as "infection of the germ," nor yet offer any apology for recording the results thus far obtained.
APPENDIX.
TELEGONY.
I.
Leaving out of consideration the question whether acquired (i.e. non-congenital) characters are transmitted, there is no problem that claims wider attention at the present time than what is now generally known as Telegony. [From tele, at a distance, and gonos, offspring.] Both naturalists and breeders are most anxious to ascertain whether what has frequently been termed the "infection of the germ" is possible - whether there is any reason for the widspread belief that the first sire influences the after progeny obtained by subsequent sires.
That the first impregnation has a lasting influence has apparently long been credited by breeders, and since Lord Morton (in 1820) communicated his famous letter to the Royal Society, many naturalists have admitted the possibility of the previous sire influencing the after progeny, i.e. leading to an extraordinary kind of reversion or "throwing back." As Lord Morton's "experiment" is so often referred to, it will be well to give in full the letter he addressed to the President of the Royal Society. The letter is as follows:
"My Dear Sir, - I yesterday had an opportunity of observing a singular fact in natural history, which you may perhaps deem not unworthy of being communicated to the Royal Society. Some years ago I was desirous of trying the experiment of domesticating the quagga, and endeavoured to procure some individuals of that species. I obtained a male; but being disappointed of a female, I tried to breed from the male quagga and a young chestnut mare of seven-eighths Arabian blood, and which had never been bred from. The result was the production of a female hybrid, now five years old, and bearing, both in her form and in her colour, very decided indications of her mixed origin. I subsequently parted with the seven-eighths Arabian mare to Sir Gore Ouseley, who has bred from her by a very fine black Arabian horse. I yesterday morning examined the produce - namely, a two-year-old filly and a year-old colt. They have the character of the Arabian breed as decidedly as can be expected where fifteen-sixteenths of the blood are Arabian, and they are fine specimens of that breed; but, both in their colour and in the hair of their manes, they have a striking resemblance to the quagga. Their colour is bay, marked more or less like the quagga in a darker tint. Both are distinguished by the dark line along the ridge of the back, the dark stripes across the fore-hand, and the dark bars across the back part of the legs. The ' stripes of the colt are confined to the withers, and to the part of the neck next to them. Those on the filly cover nearly the whole of the neck, and the back as far as the flanks. The colour of her coat on the neck adjoining to the mane is pale, and approaching to dun, rendering the stripes there more conspicuous than those on the colt. The same pale tint appears in a less degree on the rump; and in this circumstance of the dun tint also she resembles the quagga. The colt and filly were taken up from grass for my inspection, and, owing to the present state of their coats, I could not ascertain whether they bear any indications of the spots on the rump, the dark pasterns, or the narrow stripes on the forehead, with which the quagga is marked. They have no appearance of the dark line along the belly, or of the white tufts on the sides of the mane. Both their manes are black; that of the filly is short, stiff, and stands upright, and Sir Gore Ouseley's stud-groom alleged that it never was otherwise. That of the colt is long, but so stiff as to arch upwards and to hang clear of the sides of the neck, in which circumstance it resembles that of the hybrid. This is the more remarkable, as the manes of the Arabian breed hang lank, and closer to the neck than those of most others. The bars across the legs, both of the hybrid and of the colt and filly, are more strongly defined and darker than those on the legs of the quagga, which are very slightly marked; and though the hybrid has several quagga marks, which the colt and filly have not, yet the most striking, namely, the stripes on the fore-hand, are fewer and less apparent than those on the colt and filly. These circumstances may appear singular; but I think you will agree with me that they are trifles compared with the extraordinary fact of so many striking features, which do not belong to the dam, being in two successive instances communicated through her to the progeny, not only of another sire, who also has them not, but of a sire belonging probably to another species, for such we have very strong reason for supposing the quagga to be,
I am, my dear Sir,
Your faithful humble servant,
Morton.
Dr. W. H. Wollaston.
"P.S. - I have requested Sir Gore Ouseley to send me some specimens of hair from the manes of the sire, dam, colt, and filly; and I shall write to Scotland for specimens from those of the quagga and of the hybrid. I am not apt to build hypotheses in a hurry, and have no predilection either for or against the old doctrine of impressions produced by the imagination; but I can hardly suppose that the imagination could pass by the white tufts on the quagga's mane and attach itself to the coarseness of its hair.
Wimpole Street, August 12th, 1820."
NOTE BY DR. WOLLASTON.
"By the kindness of Sir Gore Ouseley, I had an opportunity of seeing the mare, the Arabian horse, the filly, and the colt, and of witnessing how correctly they agreed with the description given of them by Lord Morton." ['Philosophical Transactions,' 1820, p. 21.]
In considering this case, the question at once arises, Were the quagga-like markings of the colt and filly due to the mare having been infected by the quagga, or were they due to reversion - to the reappearance of certain ancestral characters? To this question a final answer has not yet been given. Quagga-like bands are often seen on the shoulders and legs of horses. I have before me a photograph of a Norwegian pony taken some days ago, which shows four distinct bands on the shoulder, in addition to the dorsal stripe, and distinct bands on the neck and trunk.
Darwin, after devoting much attention to the occurrence of bands on horses, states that "in all parts of the world stripes of a dark colour frequently appear along the spine, across the legs, and on the shoulders, where they are occasionally double or treble, and even sometimes on the face and body, of all breeds of horses, and of all colours." ['Animals and Plants,' vol. ii, p. 15.] Nevertheless, after discussing at some length Lord Morton's case, he concludes by saying, "there can be no doubt that the quagga affected the character of the offspring subsequently got by the black Arabian horse." [Ibid., vol. i, p. 435.]
Since Lord Morton's letter was published, numerous cases of supposed "infection of the germ" have been put on record, and quite recently a keen discussion on telegony has been carried on in the pages of the Contemporary Review between Weismann, Herbert Spencer, and Romanes.
Spencer firmly believes in telegony, and sets forth at considerable length how the germ -cells are infected. He apparently believes that as the embryo develops, germ-plasm passes from it into, and becomes a permanent part of, the body of the parent, and that later some of this germ-plasm everywhere diffused reaches and is incorporated in the germ-cells - i.e. he believes in what might be called the indirect "infection of" the germ.
Romanes, in giving the main results of a lengthened inquiry on this subject, writes as follows: ['An Examination of Weismannism,' 1893, Appendix II.] "The investigations have been pursued on three different lines: (1) I raised discussions on the subject in the principal breeders' and fanciers' journals of this country and also of America. (2) I entered into private correspondence with contributors of the largest experience, and also with professional and amateur breeders, fanciers, &c., who addressed me directly on the subject. (3) I started experiments with the varieties which these inquiries indicated as most likely to yield positive results. At present nothing need be said with regard to these experiments, because they are not sufficiently matured; but it is desirable to state the general upshot of the correspondence.
"The principal result is to show that the phenomenon is of much less frequent occurrence than is generally supposed. Indeed, it is so rare that I doubt whether it takes place in more than one or two per cent, of cases. I must add, however, that nearly all my professional correspondents would deem this an absurdly low estimate. It has hitherto puzzled me why the phenomenon in question, since it does certainly occur in some cases, should occur so rarely. But I think that Mr. Spencer's suggestion on this point is a valuable one. This suggestion is that when the first sire is of a relatively stable and also of a markedly different ancestral stock from the dam, there will be most likelihood of his impressing his ancestral characters on the progeny of the second sire."
While Romanes evidently believes in the possibility of telegony, he is far from accepting Spencer's explanation of the phenomenon. He adopts the simpler explanation that the unused germ-plasm from the first sire directly infects - gains access to - the unripe ova. In holding this view, Romanes seems to be in agreement with Darwin, who apparently believed that the male element acts directly on the reproductive organs of the female, and not through the intervention of the crossed embryo. Another believer in telegony who may be mentioned is Agassiz. As the result of various experiments, he satisfied himself "that the act of fecundation is not an act which is limited in its effect, but that it is an act which affects the whole system, the sexual system especially; and in the sexual system the ovary to be impregnated hereafter is so modified by the first act, that later impregnations do not efface that first impression."
But while Darwin, Spencer, Agassiz, and Romanes believe more or less firmly in telegony, there are many who are either doubters or unbelievers. The most prominent of these is Weismann, who inclines to the view that there is no such thing as an "infection of the germ." In a recent number of the Contemporary Review [Vol. Ixiv.] he writes as follows:
"I must say that to this day, and in spite of the additional cases brought forward by Spencer and Romanes, I do not consider that telegony has been proved. I do not dispute the possibility of telegony; I grant that the wide general acceptance of the belief in the past has so impressed me that I have always said that possibly it might be justifiable and founded on fact. I should accept a case like that of Lord Morton's mare as satisfactory evidence if it were quite certainly beyond doubt. But that is by no means the case, as Settegast has abundantly proved."
Weismann does not doubt that after the mare had borne a hybrid to a quagga she subsequently had colts by a horse, and that these were marked with stripes on the neck, withers, and legs, but he contends that there were no other characteristics of the quagga discernible in the colts. The stripes do not in themselves, Settegast thinks, amount to proof, "for every experienced horse-breeder knows" that "cases are not very rare in which colts are born with stripes that recall those of the quagga or zebra. They regularly disappear as the colts increase in age." Weismann refers to other experiments, and adds, "the attempt must be made to determine the truth by new experiments."
"But as hitherto," Weismann adds, "there have been no positive results from the observations that have been made; and as the most competent judges, namely, breeders who have a scientific knowledge, such as Settegast and Nathusius, and the late head of the Prussian Agricultural Station at Halle, Professor Kuehn, spite of their extensive experience in breeding and crossing, have never known a case of telegony, and therefore have great doubts as to its reality, it seems to me that, ‘according to scientific principles, only the confirmation of the tradition by methodical investigation, in this case by experiment, could raise telegony to the rank of a fact.' "
In concluding his observations on telegony," ['Germ-plasm,' p. 3S5.] Weismann says: - "Experienced breeders, like Settegast and Kuehn, of Halle, do not believe in it, for though they have crossed various domestic animals they have never observed an instance of it. . . . If the 'infection' were proved beyond a doubt, a supplementary fertilisation of an egg-cell in this manner must be considered possible; we certainly might then reasonably ask why mares, cows, or sheep should not occasionally become pregnant without being served a second time. But this has never yet been known to occur, and I incline to Settegast's view, that there is no such thing as an 'infection' of this kind, and that all the instances which have been recorded and discussed critically by him are based upon a misconception."
Thus the case stands at the present day. At the one extreme we have Weismann and certain experienced German breeders; at the other many English breeders so persuaded that telegony is of frequent occurrence that they make a point of always mating their mares, &c., first with a male having a good pedigree, so that their subsequent progeny may be benefited by his influence even though inferior sires are subsequently resorted to.
Evidently this is a question of as much practical im- portance as it is of profound scientific interest, and it is a question which both men of science and breeders are alike agreed should, if possible, be settled without further delay. By way of making an attempt in this direction I am anxious to organise an extensive series of experiments, and my chief object in writing this paper is to enlist the interest and co-operation of breeders and others in a position to render assistance.
I propose to repeat as exactly as possible Lord Morton's experiment, using, however, a zebra in place of the now extinct quagga. I have already obtained a number of mares, but as, according to Romanes, telegony may only occur in a very small percentage of cases, I shall be glad to hear from any one who is willing to place a suitable mare at my disposal. But in addition to crossing the horse and zebra, it is most desirable that experiments be made with cattle, sheep, pigs, dogs, &c., of various breeds. May I venture to appeal to those sufficiently interested in the question who happen to be in a position that admits of their taking part in the work to communicate with me, so that the necessary arrangements for further experiment may be made. In the next number of this Journal I shall give a list of the experiments which I think might be started during the present season.
II.
In the paper on Telegony published in the April number of the Veterinarian, after quoting in full the now famous letter of Lord Morton, I especially referred to the views of Darwin, Herbert Spencer, Agassiz, Romanes, and Weismann. Since this paper appeared I have had the opportunity of reading the recently published work on horses, asses, &c., by Tegetmeier and Sutherland. [‘Horses, Asses, Zebras, and Mules,' Horace Cox, Field Office, 1895.] As the one writer has long taken a keen interest in almost all branches of natural history, and more especially in the crossing of various kinds of animals, while the other has been long successfully engaged in breeding mules, it will be well to indicate their views on the subject under consideration. They better than any others may, I think, be held as representing the views generally prevailing amongst not only breeders, but all who take an interest in our domestic and semi-domestic animals.
Some breeders of mules accept the infection of the germ theory; others, who have had considerable experience in the United States, affirm they have never seen any evidence of the influence of the first sire on the subsequent progeny to other sires.
Believers in telegony frequently refer to the progeny of an African ass [Equus asinus) obtained by the Zoological Society of London in 1881. This ass produced her first foal in June, 1883, the other parent being an Asiatic ass (E. hemiouus). The hybrid foal closely resembled the male parent, which was reddish in colour, the female being of the usual grey. In 1889 and again in 1891 this female African ass produced a foal to an African male ass. Both foals presented the usual characters of the common or African ass. But in 1892 a third foal was born to the same African male ass; and this foal was of a reddish fawn-colour, somewhat shaggy in coat, and provided with a large star on the forehead and a white blaze down the face, while the ears were relatively short and the shoulder and leg stripes indistinct. [Recently (1898) this ass has produced a second reddish-coloured foal like her first mate the Asiatic ass.]
This departure from the normal type might either be due (1) to simple reversion, (2) to what would now-a-days be designated discontinuous variation, or (3) to telegony - in this case to the influence of the Asiatic first sire.
In discussing this case the writers say: "The influence of the first sire on all subsequent offspring is a subject of very considerable importance, that has not received the scientific investigation that it merits. It is generally accepted by breeders of dogs, and in the case of valuable animals the effect of a mesalliance is carefully guarded against. It is one that is recognised by physiologists as affecting the human species, and the example of the striped foals that were always bred by a mare whose first foal was a hybrid of zebra parentage is well known. [I have never heard of a well-authenticated case of telegony in the human species - J. C E.] Whether this young ass bred in the Gardens is merely an accidental variation, or whether it owes its peculiarities to the influence of the Hemippe [Asiatic first sire] is a point I will not endeavour to decide." [Ibid., pp. 15 and 16.]
In the chapters on mules and mule-breeding the influence of the first sire is again referred to. In support of the statement that male and female mules and hinnies are, as far as is known, "absolutely sterile," the authors quote from Captain Hayes as follows: "Neither the mule (the produce of the jackass and mare) nor the hinny or jennet (the cross between the horse and the she-ass) is fertile, either among themselves or with other members of the horse family. Those animals which have been mistaken by superficial observers as fertile mules have been, I venture to say, in most cases the off- spring of mares that have previously bred to donkeys, and have endowed their young with some of the characteristics of their former asinine lovers. Both the mule and the jennet respectively 'take after their dam in size, and their sire in appearance and disposition."
That the authors agree with Captain Hayes may be gathered from what they say as to the "mule" which produced foals to both the horse and the ass in the Acclimatisation Gardens in Paris. After pointing out that "it is doubtful whether the animal is a mule," they add, "It is not at all improbable that her female parent had bred a mule in the first instance, and, as in the well-known cases of mares which have been mated with quaggas and zebras, her subsequent progeny, when mated with a horse, shows some trace of the first union. The late M. Ayrault, and most persons who are really cognizant of the matter, regard this animal not as a mule but as an ordinary mare. ... It would appear most probable that this is not a case of a fertile mule breeding; but that the animal is really an ordinary mare, whose female parent was influenced by a first alliance, as is so often the case in dogs and other animals." [Ibid., pp. 81 and 82.]
From these quotations it may be assumed (1) that Tegetmeier and Sutherland believe in telegony in "the case of dogs and other animals," and that it most probably occurs also in horses, and (2) that Captain Hayes (who probably expresses the views of the majority of veterinarians) takes for granted that telegony occurs in horses as well as in other animals.
The views held, by biologists, breeders, and others interested in the question of telegony having been shortly stated, I shall now indicate some experiments that might at the outset be started. Granting that infection of the germ is possible, various questions at once arise. Is it, as believed by Romanes, extremely rare, or is it, as asserted by many breeders, of common occurrence? It is conceivable that it may be of common occurrence, but owing to inbreeding and other causes the results in the majority of cases may be completely or all but completely obscured. In planning experiments it will, I think, be well to take for granted that if telegony occurs at all it occurs frequently, and that in order to demonstrate the influence of the first sire it is only necessary to select suitable subjects.
Judging by the cases already reported which seem to support the belief in telegony, it will be desirable to, in the first instance, operate with either different species or well-marked varieties of the same species, and then mate the females which have bred to the new species or distinct varieties with males of their own particular breed. The following are the crosses, that I think might well be tried.
1. A male zebra and various breeds of mares, - each fertile mare to be afterwards mated with an Arab horse, or a horse of its own particular breed.
2. An Arab or other distinct breed of horses and a zebra mare, the zebra mare, if fertile, to be afterwards mated with a male zebra.
3. A male zebra and a jennet, the jennet to be afterwards bred to a jackass.
4. A jackass and a mare, the mare to be afterwards mated with a horse.
5. A zebu bull and a Kerry and an Ayrshire cow, the cows to be afterwards bred to their respective bulls.
6. A Chillingham bull and Galloway, Angus, Norfolk, and Devon cows, each cow to be afterwards mated with a bull of its own breed.
7. A Galloway bull and a shorthorn cow, the cow to be afterwards mated with a shorthorn bull.
8,. An Aberdeen Angus bull and a shorthorn cow, the cow to be afterwards mated with a shorthorn bull.
9. A red shorthorn and a Dutch cow, the cow to be afterwards mated with a Dutch bull.
10. A Leicester tup and a black-faced ewe, the ewe to be afterwards mated with a black-faced tup.
11. An Iceland tup and a Shropshire ewe, the ewe to be afterwards mated with a Shropshire tup.
12. A Southdown tup and a merino ewe, the ewe to be afterwards mated with a merino tup.
13. A goat and a ewe, the ewe, if fertile, to be afterwards mated with a tup of her own breed.
14. A Berkshire boar and a Tamworth sow, the sow to be afterwards mated with a Tamworth boar.
15. An Irish boar and a Tamworth sow, the sow to be afterwards mated with a Tamworth boar.
The above are merely offered as suggestions; many other crosses might be equally or even more successful in throwing light on the question at issue. I shall be glad to hear from anyone willing to undertake any of the above or similar experiments. I have not made any suggestions as to experiments with dogs, rabbits, ducks, fowls, &c. This is not because I think valuable results might not be obtained, but rather because breeders and fanciers willing to take part in the investigation, can settle best for themselves, on what particular lines they might most successfully work with the domestic animals not specially referred to. The first three crosses suggested I am making arrangements to carry out with as little delay as possible.
III - ORIGINAL TEXT OF ARTICLE IN "THE ZOOLOGIST".
ON ZEBRA-HORSE HYBRIDS.
By J.C. Ewart, F.R.S., Regius Professor of Natural History, University of Edinburgh.
The Zoologist, No. 680.: - February, 1898.
During the last two years I have bred five hybrids by crossing mares with a Zebra (Equus burchelli var. chapmani). The first hybrid was born on Aug. 12th, 1896; the others were born during the summer of 1897. The dams of the respective hybrids are (1) an Island of Rum pony, (2) a Shetland pony, (3) an Iceland pony, (4) an Irish mare, (5) a cross-bred Clydesdale mare.
The Sire “Matopo”
The sire ("Matopo") of all the hybrids is a handsome 12.3 hands Burchell's Zebra, probably from the Transvaal. As fig. 1 (PI. I.) shows, Matopo is well formed, with powerful legs and, for a Zebra, a fine neck and fairly good shoulders. In his movements he is almost perfect. When trotting, the fore legs move gracefully, without suggesting the hammering action of the hackney; and when galloping he seems to bound along as if without effort, and with but little expenditure of energy.
If Zebras deserve the ill character they have hitherto borne, Matopo must be an exception to the rule. We are too apt to forget that until Zebras have been under domestication for some generations, it is unfair to judge them by the Horse standard, which after all is not so very high. I have known several perfectly docile captured Zebras, and I have had in my possession a filly (taken when quite young in the Transvaal) which from the first was as docile, tractable, and trustworthy as any pony that was ever foaled. I have refrained from handling Matopo for obvious reasons, yet there is never any difficulty in managing him, unless when he is herding mares, or unusually excited. When in a field with mares he is unapproachable, for, regardless of consequences, he attacks all who venture into his vicinity. Galloping up open-mouthed, uttering his characteristic call, he endeavours to seize intruders by the legs. On one occasion, in a small paddock, he guarded a dozen mares so well, that it took four of us nearly two hours to drive them into their boxes. He is, however, easily upset by unusual noises, and there is nothing that drives him into a state of frenzy so readily as carpet-beating, or that cows him so effectually as a coil of rope. I have often wondered if the rhythmic beating of carpets reminds him of the day when in far-off Africa he lost his freedom; of the time when Boers entangled his limbs to music made by Zulus beating their shields with their assegais.
The more characteristic stripes of Matopo are seen in figs. 1 and 2 (Pl. I.). I have already described at some length ['Veterinarian,' November, 1897] the plan of the striping in various Zebras, and hence only a short account of Matopo's markings need here be given.
Fig. 2 (Pl. I.) shows a series of pointed brow arches, some of which end in a frontal tuft nearly two inches in length. Continuous with the frontal stripes are a number of vertical stripes. These stripes extend to the muzzle, the dark skin of which is sparsely covered with short light hairs, except above the nostrils where there are dark brown "nostril patches." There is usually a distinct shoulder-stripe in Zebras, passing downwards from the withers to bifurcate about the level of the shoulder-joint. In fig. 2 (Pl. I.) the shoulder-stripe is double, while in fig. 1 it has blended with a humeral stripe. Between the shoulder-stripe and the occipital crest there are usually twelve cervical stripes, all of which run up into the mane to form, with a corresponding number of white bundles, a series of black and white tufts. Lying between the two upright rows of tufts, and continuous with the dorsal band, is the mane proper, consisting of more or less upright black hairs. The most anterior part of the mane, instead of forming a forelock, extends beyond the level of the ears, and projects forwards at right angles to the long axis of the face. Behind the shoulder-stripe there are, on the left side, five broad, nearly vertical stripes, all but the last reaching the dorsal band above, while all but the first reach the ventral band below. Behind the fifth vertical stripe are a number of broad oblique stripes, with indistinct "shadow" stripes between them. One of these oblique stripes, beginning at the root of the tail, runs forward to pass over the point of the ilium (hip) before bending sharply downwards to reach the ventral band. I have named this the great flank-stripe. Below this flank-stripe a second, having a similar course, may be known as the intermediate flank-stripe. The intermediate stripe is followed by a third, which, starting some distance below the root of the tail, runs obliquely across the quarters to bifurcate over the stifle, the anterior division proceeding towards, but not actually reaching, the ventral band. This may be known as the lower or stifle flank-stripe. These three flank-stripes are equally distinct on the right side, the bifurcation over the stifle being especially evident.
In the space formed by the splitting of the shoulder-stripe are several indistinct arches, and below these arches are the transverse bars of the leg. In some cases this V-shaped space contains portions of seven arches, and the legs may be striped to the hoofs. Below the stifle-stripe there are first oblique and then nearly transverse stripes across the hind leg, with sometimes shadow-stripes between. In Matopo the stripes are indistinct on the lower part of the hind leg, but in many Zebras they become more distinct and relatively broader as the hoof is reached. It will be further observed from the figures (a) that the upper part of the tail is distinctly striped, and that, as in an Ox, only the lower part of the tail carries long hairs; (b) that though there is a large wart (chestnut) on the fore leg, there is no vestige of a wart on the hind leg, and (c) that there is no tuft of hair at the fetlock.
It may be mentioned that in no two Zebras, or on the two sides (Plate I., figs. 1 and 2) of the same Zebra, is the striping alike, that in some cases there are nearly as many shadow as there are ordinary stripes on the neck and body; that even in some Burchell's Zebras there are stripes across the croup and rump which suggest the "gridiron" of the Common Zebra (E. zebra); and that while in summer the dark stripes are nearly black and the light stripes cream-coloured, in winter the dark stripes are occupied by fairly long brown hairs, while the light stripes are made up of equally long white hairs; the light tufts at the side of the mane, however, are white summer and winter. It may be added that Matopo, like the majority of the Burchell group of Zebras, being adapted for a life on the plains, has rounded hoofs and comparatively short ears. He thus differs from the Mountain Zebra (E. zebra), and from his stable companion, a white Egyptian Donkey, in which the hoofs are long and narrow, while the ears measure 11½ in., five inches more than in Matopo.
The Hybrid "Romulus."
The oldest hybrid (Romulus), as already noted, was born on the 12th of August, 1896, the period of gestation being three hundred and forty-two days,: - in the mare it is usually from three hundred and forty to three hundred and fifty days. The dam of Romulus was a 13-hands, black Island of Rum pony, lent for the experiments by Lord Arthur Cecil, of Orchardmains, Kent. The well-bred black ponies of the Scottish Western Highlands and Islands, which have long been under observation, form a distinct breed, well adapted in many ways for crossing with Zebras. Their resemblance to Eastern Horses has been accounted for by saying they have descended from sires which escaped from the ships of the Spanish Armada. [Further particulars as to Mulatto, the dam of Romulus, will be found in the 'Veterinarian' for November, 1896]
Romulus, when a few days old, was the most attractive little creature I have ever seen (Plate III., fig. 1). He seemed to combine all the grace and beauty of an Antelope and a well-bred Arab foal. Instead of, like his sire, looking as if freshly painted for a Lord Mayor's Show, he was faultless in colouring and in the disposition of the stripes, spots, and bands. The body colour was chiefly of a bright golden yellow, while the stripes and spots were of a rich dark brown; but what was especially remarkable was the indescribable sheen of his coat, the dark bands being especially lustrous. A casual glance showed that in the plan of his striping Romulus was utterly unlike his sire, and, when a careful examination was made, it became evident that in the number and arrangement of the markings he was not unlike a Somali Zebra. As fig. 1 (PI. II.) shows, the brow has been tattooed as if to represent a huge finger print. Instead of the four or five acutely-pointed frontal arches of his sire, there are fourteen rounded arches, that remind one of the face of the Somali Zebra. Instead of twelve cervical stripes, as in Matopo, there are in Romulus twenty-four cervical stripes, all of which can be traced into the mane. In having so many cervical stripes, he seems to be more primitive than even the Somali Zebra (in which I have never seen more than fourteen cervical stripes), but closely agrees with one of my Zebra mares when the shadow stripes are included. The shoulder stripe bifurcates higher up than in Matopo, and there are seven indistinct arches in the triangular space below the point of bifurcation. Behind the shoulder stripe there are nine (Plate III., fig. 1) fairly distinct vertical stripes instead of five, as in his sire (Plate I., fig. 1). Apparently corresponding to the three flank stripes so often seen in Burchell Zebras, there are in the hybrid three stripes in front of the stifle, which first run upwards and then arch backwards to end below the root of the tail (Plate III., fig. 1). In the triangular space between the first flank stripe and the ninth vertical body stripe are numerous narrow indistinct lines, some of which proceed towards the ventral band, while others join the first or great flank stripe. In line with these nearly transverse stripes there were at birth numerous spots arranged in nearly transverse rows over the loins and rump. Now that the hybrid is over a year old (Plate III., fig. 2) most of the spots have united to form somewhat zigzag narrow bands, almost identical in their direction with the narrow stripes over the hind quarters of the Somali Zebra. On the left side the blending of the spots has advanced further than on the right. Counting from the shoulder stripe to the root of the tail there are forty-three stripes in the hybrid,: - about the same number as in the Somali Zebra; in Matopo there are only five transverse stripes behind the shoulder stripe (Plate I., fig. 1). It seems to me the blending of the spots over the hind quarters of Romulus goes a long way towards proving that stripes are in many cases first represented by spots or interrupted zigzag wavy line?. Between the stifle or third flank stripe and the point of the hock there are a number of dark bands (between some of which are shadow stripes), while below the hock there are first several distinct transverse bars, and then a number of less distinct oblique lines, right down to the hoof. Similar bars and lines occur on the fore-limb. These leg bars were at birth more distinct than in the Zebra sire. Continuous with the mane is a well-defined dorsal band (with a narrow yellow band at each side) which extends some distance into the tail. The tail in the hybrid had, at birth, long hairs right up to the root, but, notwithstanding this, there were three distinct bars visible at each side; similar tail bars I have once seen in a Horse.
Though the ears look long in some of the photographs, they are now relatively very little longer (though rounder at the apex) than in the majority of Horses. The nostrils, in their shape, position, &c, are Zebra-like, and the eyes and eyebrows may be said to be intermediate; but the eyelashes are long and curved, and quite unlike the short almost straight eyelashes of Zebras and Horses. The feet of Romulus suggest the Zebra more than the Horse. They seem to be made of excellent stuff, and to stand a good deal of wear. In his movements, the hybrid takes more after his sire than his dam. A few minutes after birth he was rushing about his box, impatient apparently to join the parental troop. What has struck me from the first has been his alertness and the expedition with which he escapes from suspicious or unfamiliar objects. When quite young, if caught napping in the paddock, the facility with which he, as it were, rolled on to his feet and darted off was wonderful. The principal enemy of the Zebra seems to be the Lion. To escape from the Lion, great and sustained speed is not so requisite as a decided and rapid bound when the Lion makes his spring, or when he is accidentally met with in the veld. This rapidity of getting out of the way has been strongly inherited by all the hybrids. Zebras, as far as my experience goes, are difficult to handle, not so much because they are vicious or intractable, as because they are afraid. At any moment they may be seized by panic,: - when they imagine there is a Lion in the path,: - and, regardless of consequences, rush, it may be, against a wall or a hedge, or into a ditch, reins and bits counting for little or nothing. In schooling the hybrids, this habit will require to be allowed for, and the tendency to bound or rush slowly combated. As it has been completely overcome by careful training in some Zebras, there should be comparatively little difficulty in breaking the hybrids. As a matter of fact. Romulus leads anywhere, is perfectly docile, allows his feet to be trimmed and his teeth to be examined, and, when little more than a year old, seemed quite willing to carry a small boy on his back.
I mentioned Mulatto is just under 13 hands, while the Zebra sire is nearly 12.3 hands. At birth (August 12th, 1896) Romulus measured 34½ in. (from the withers to the ground); at two months 38½ in.; at six months 43 in.; and at twelve months 45½ in. The rate of growth has been extremely inconstant,: - e.g. from the 12th of February to the 12th of April he only increased half an inch, [He was weaned on the 14th of February, and fretted not a little for some time after.] and from the 12th of June, 1897, to the 12th of September, 1897, he only increased three-quarters of an inch [During the greater part of this period he was shedding his old and growing a new coat.]; but from the 12th of September, 1897, to the 12th of December, 1897, he increased one and a quarter inches. He now measures (January 12th) 47½ in., nearly 12 hands, and the circumference of the fore-shank is 6⅛ in., the knee being 10 in., and the girth 52½ in.
The foals of the black Island of Rum ponies are frequently of a mouse-dun colour, with at times an indistinct dorsal band, and a cloudy patch over the shoulder. Usually after the first coat is shed the pure-bred foals are dark brown, and later nearly black, with sometimes indistinct dappling over the flanks and hind quarters. As already mentioned, the body colour of Romulus at birth was chiefly of a yellow tint, the yellow approaching bright orange on the brow, while it approached a straw colour at the muzzle and below the knees and hocks. Under the neck and under the belly the prevailing body colour was dark brown, the ventral band being very indistinct.
The ears were lined with fine bright orange-coloured hairs. When only a month old, the hybrid began to shed his foal's coat. The light-coloured hairs began to drop out from the face and neck about the middle of September, and by the end of September he looked considerably darker. The yellow and also the dark brown hairs continued to fall out, except over the back, all through October, and by the middle of November only the orange-coloured lining of the ears was left to remind one of the rich coat he wore during the earlier weeks of his life. By the end of November the new coat was established. The bright orange facial bands were replaced by much paler bands, the muzzle was nearly brown in colour, the neck and body intermediate spaces approached a mouse-dun colour, while the lower parts of the legs were of a dark brown tint. From the withers to near the root of the tail the hair was especially long and thick. For a time the hair over the croup and the greater part of the rump was so much longer than the hair around the root of the tail that it looked as if part of the hind quarters had been previously clipped. The new coat consisted of a thick layer of woolly hair, from half an inch to nearly two inches in length, and of a less complete coat of stronger hairs, many of which were nearly three inches in length. Near their roots all the body hairs were light in colour, which implies that had the hybrid been clipped, there would have been little or no indication of stripes left. In the Zebra, on the other hand, the dark pigment extends to the roots of the hair, and hence, however short the hair may be, the banding is quite evident. Recently the skin around the root of Matopo's tail was injured, with the result that the hair, together with some of the epidermis, was shed; but even before the points of the new hair could be detected, the position of the dark bands was perfectly distinct. The skin of the Zebra has been described as uniformly black, even under the white bands; but it would be more accurate to say it is of a nearly uniform dark grey colour.
About the middle of March the long hairs began to drop out, and by the end of March they came away in handfuls. As the long hairs were shed from the body, the long hairs were shed from the upper half of the tail, with the result that for a time the tail of the hybrid was little better covered than the tail of his sire. By the end of May all the long hairs: - light and dark: - had vanished, and early in June the dark and mouse-coloured woolly hairs were coming out. By the 6th of June the dark lustreless winter coat had sufficiently gone around the base of the ears and above the eyes to indicate the colour of the summer coat. All through June and July the process of shedding continued, but by the 12th of August: - the hybrid's first birthday: - the summer coat was fully established. The dark stripes, which consisted chiefly of strong flattened hairs, looked very prominent. The intermediate bands were of a reddish brown colour over the brow, but elsewhere reminded one of the summer coat of a Stag. Taken as a whole Romulus was very decidedly darker as a yearling than during the early weeks of his existence.
As the long hairs were shed from the body and the root of the tail, numerous hairs dropped from the mane. In an ordinary mule (the foal of a New Forest pony) which I have had for some time, all the long hairs of the mane were shed last summer; but in Romulus, either some of the long hairs were retained, or the new hairs came in before the old ones were lost. At any rate, though the mane was shorter and less bulky and consequently more upright during August, it always consisted of numerous long hairs. At present the mane, which consists of wavy hairs from seven to nine inches in length, tends to fall slightly to one side,: - the mane falls slightly to one side in some Zebras.
By the middle of September Romulus had again lost not a few of the brighter coloured hairs, and since then he has been getting again gradually darker. Probably because of the extreme mildness of the season the long hairs have already (January) begun to fall out in much the same way as they did last March.
All the experts who have seen Romulus agree in considering him a decided improvement on his sire, and more attractive and shapely than his dam. Having been handled from the first, he is, as a rule, extremely quiet. Occasionally, however, he clearly indicates he has plenty of courage and no lack of speed. At present he is particularly attached to a small thoroughbred mare. When separated from this mare he is sometimes as restless as his sire when upset by some change in his surroundings. Last week a strange Horse was galloped in the paddock where Romulus happened to be for the day. The hybrid became excited, and gave an excellent demonstration of his trotting and galloping powers, and of how proudly he could carry himself, and this continued for some time after the intruder left the field. Romulus was recently described by an excellent judge of Horses in the 'Scottish Farmer' as "a bonnie colt, with rare quality of bone, ... and with the dainty step and dignity of the Zebra." There is nothing about the hybrids, strange to say, that suggests the ordinary mule or hinny.
The Hybrid "Remus."
The dam of Remus is a three-parts bred, 14.1-hands Irish mare. "Biddy" has been in my possession since 1893, and is now nine years old. She is a bay, with black points, but no white hairs anywhere, and Remus is her first foal. She is a very gentle quiet creature, and has always been in excellent condition, winter and summer alike.
Evidently the Zebra, before coming here, had not made the acquaintance of any of his equine relatives. When first introduced to Mulatto, he rushed into a corner with his tail between his legs, and uttered peculiar little sounds which strongly suggested abject fear. Some of the ponies rushed at him open-mouthed; others deliberately pelted him with their heels. On the other hand, a bay Arab stallion and various mares could not have been more alarmed had he been a Tiger, or, when he called "Quacha," "Quacha," a troup of Lions. To give him a chance of discovering what sort of an animal a Horse is, I turned him loose one evening with a good-natured but very plucky bay Shetland pony. The pony proceeded to tease the Zebra, who very soon began to show fight. He was soon circling round the pony with the object of seizing her legs. For a time the pony was unprepared for this mode of attack, but ere long adopted similar tactics, with the result that the Zebra was several times brought to his knees. [I may mention that when his legs are touched with a rope or stick he almost invariably drops on to his knees, or lies down altogether. This is, I think, the result of his having been periodically thrown before he came here that his hoofs might be looked to.] After a couple of hours the duel came to an end,: - the damage being very slight on either side,: - and ever afterwards Matopo and "Sheila" were excellent friends. But even during the spring of 1896 the Zebra was ridiculously timid, and even now a very small demonstration leads him to beat a hasty retreat. Biddy was the first fairly large animal he ventured to approach. One day I tied her up in a court about forty feet square, a cloth having been previously bound over her eyes. The Zebra in course of time ventured within a few yards; later he laid his head across her quarters, and then, for quite a long time, across her withers. He next licked her lips, and ended by gently nibbling at her ears. Evidently at length satisfied a big Horse was after all not so terrible an object, he retired to his box and finished his corn. Having once learned the peculiarities of a mare he never forgets them. Some of the mares he dislikes, while he is very fond of others, getting quite excited when they pass his own particular quarters. Donkeys, however, he completely refuses to take the smallest notice of.
Remus: - born on the 18th of May, 1896: - was, at birth, relatively smaller and far less active than Romulus; the period of gestation was three hundred and forty-six days. When a day old he measured 35½ in., his girth being 28 in. On the 18th of June he had increased to 38¾ in., the girth being 36 in. When six months old he measured 44⅞ in., the girth being 47½ in., the circumference at the knee 9¾ in., and below the knee 5¾ in. Romulus at six months was 42 in.
From the first Remus has been extremely friendly, and yet in some respects he is more Zebra-like than Romulus. For some days he was little more than a machine,: - an automaton capable of following a moving object and of sucking. All the special sense organs were apparently at work, but the brain seemed incapable of making much use of the information collected. If I moved away he followed me, and sucked at my fingers or anything else offered him. He heard his dam when she called, but he was unable to discover whence the sound came, and when he saw her at a few yards distance he failed to recognize her. He seemed to like aloes and water quite as much as sugar and milk, and did not mind either strong smelling-salts or freshly-made mustard. Though he kicked aimlessly when pinched, he paid no heed to the application of either warm or very cold substances to his skin. When a dog was first introduced to Romulus, his excitement was intense. He rushed about at a furious rate, striking as opportunity offered with his fore-feet, and holding his head high and stepping high, as if moving through long grass, where other enemies might lie concealed. Remus, on the other hand, when two days old, allowed a yellow collie to lie down within six inches of his muzzle, and only got up as a Dalmatian approached when a warning note was uttered by his watchful parent. When the four hybrids and two pure-bred foals were eventually weaned, Remus seemed to mind very little. While one of the hybrids and a half Arab foal were biting and kicking and rushing about as if demented, Remus simply stood looking over the fence. But by-and-by, when the others settled down, he set to walking backwards and forwards behind the wall of his court, exactly like his Zebra sire, and though he still keeps this up as if he were a caged Lion, none of the others have followed his example. When Romulus was weaned, he for some days rushed about, as much as a Zebra when highly excited, as his sire when upset by the beating of carpets. Recently it was necessary to give the hybrids milk containing thymol. The pure-bred foals offered but little resistance, but all the hybrids fought till they were exhausted, and nothing would persuade Remus to swallow the first dose.
As might have been expected, Biddy's foal is much lighter in colour than Mulatto's. With the exception of the muzzle and the lower part of the legs, the body colour is a rich light bay; the muzzle and legs were, at birth, more of a mealy colour, but are now of a bay colour. The bands are much lighter, and consequently less distinct than in Romulus. As a rule they are of a dark reddish brown hue, being especially evident on the brow, the forearms, and above and below the hocks. The plan of the striping is the same as in Romulus; but even at birth several of the rows of spots across the croup had already united to form narrow bands. The face, measured from the occipital crest to a line connecting the upper margins of the nostrils, was slightly longer than in Romulus; but the ears were the same length: - six inches.
Sometimes when a Horse utters a warning call all the members of the herd hurriedly collect together and rush about in an excited manner. It seems to be of the utmost importance for wild Equidæ to at once make out the direction of any given sound. Probably the longer the ears the quicker this is accomplished. If the length of the ears, as is most probable, counts for much, one can understand why they almost reach their full size at birth. Foals are given to straying in all directions, and unless they hear and at once recognize the call of their respective dams, and the direction from which the sound comes, their chances of surviving in a wild state would be greatly reduced. At birth, the ears of Romulus were longer than in his dam, and only slightly shorter than in his sire. In the case of Remus they were the same length as in his dam, viz. six inches along the inner aspect.
The eyes in Biddy's foal are hazel-coloured and gazelle-like in their mildness, and the eyelashes are particularly long and curved. The mane was at first made up of soft hairs, which bent over to the right side. The mane, however, soon assumed an upright position, and now, when nearly eight months old, it consists of nearly erect but not very stiff hairs. It looks as if the mane will always be as upright and as short as in his sire. The tail contains fewer hairs than any of the other hybrids, and has three bars across the root. On the other hand, unlike ordinary Mules, there are chestnuts on the hind legs as well as on the fore. The front chestnuts are large, level with the skin, and Zebra-like; the hind chestnuts are raised above the level of the skin, and, though narrow and only half an inch in length, are Horse-like. That the Zebras and Asses have no chestnuts on the hind legs may perhaps be due to the absence of chestnuts in their remote ancestors; their absence points, I think, to Asses and Zebras having sprung from a different ancestor (perhaps Hipparion) than the Horses, which may have descended straight from Protohippus. If Remus survives, he may reach a height of nearly 14 hands, and be the most handsome and fleetest of all the present crop of hybrids.
As in the case of Zebra foals, the hair over the back and hind quarters of Remus soon increased in length, and formed a thick woolly covering. The hair of the first coat usually falls off soonest from the face and neck, then from the legs, especially at the knees and above and below the hocks. Some of the hair was shed from the face by the end of the first month, but there was still some left on the muzzle and brow at the end of the third month, and the legs retained some of the foal's coat at the end of the fourth month. The second coat, which was completed by the end of the fifth month (i.e. about the middle of October), consists of a thick inner coat of bay and brown fine wavy hairs, averaging an inch and a half in length, and of an outer but much less abundant coat of stronger hairs, many of which are 2½ in. in length. Neither the long nor short hairs nor the hairs of the mane have yet (January) begun to fall out.
The Hybrid "Brenda."
The dam ("Lady Douglas") of Brenda is a cross-bred Clydesdale mare, built on the lines of the "Douglas" breed, once common in the Hamilton district. Like Biddy, she is a bay with black points, but, unlike the Irish mare, she has a large "blaze" on the face, a heavy mane and tail, and a liberal amount of hair at the fetlock joints. Lady Douglas is 15 hands high, the circumference at the knee is 13½ in., and below the knee 9 in. The face is longer than in Biddy by nearly an inch, and the ears by three-quarters of an inch. I expected Brenda (the Clydesdale's first foal) to closely resemble Remus in colour and markings, but in breeding, more especially in cross-breeding, the unexpected often happens. We are too apt to forget that, even when the sire belongs to a different and very distinct species, the progeny may take after the cross-bred dam. It was evident soon after Brenda (Plate II., fig. 2) was foaled that she differed not a little both from Romulus and Remus. In the first place her ears looked extremely long; they were at birth 6½ in., only a quarter of an inch shorter than the ears of her dam, and quite as long as the ears of her sire. The ears now measure seven and a half inches; on the other hand the head is relatively short: - shorter than the head of a 12-hands Iceland pony's hybrid. The height at the withers was 43 in., one inch more than in Remus, and four inches more than in the Iceland hybrid. At birth Brenda, apart from her ears, looked not unlike an ordinary bay foal, but soon faint stripes began to show themselves, and in a day or two the stripes, though indistinct, were seen to closely agree in their arrangement with those of the other hybrids. Now that the "Clydesdale" hybrid is nearly seven months old, she at a little distance might easily be mistaken for an ordinary foal. Compared with Remus the head is shorter and finer, while the joints are larger and the shanks thicker. At six months the circumference at the knee was 10¼ in., and below the knee 6⅛ in.: - almost exactly the same as in Romulus when seventeen months old. The mane, at first nearly upright, short and Zebra-like, is now made up of hairs from eight to ten inches in length (nearly as long as in an ordinary foal of the same age). Except near the withers and between the ears the mane arches freely to the right side, some of the hairs almost touching the neck. The hair between the ears already projects forwards to form a forelock. In Remus, as already mentioned, the mane is still upright, and shorter than in his sire. The tail in Brenda has also from the first been heavier than in any other of the hybrids, and fewer hairs have been shed from its base; further, almost from the first there have been a few hairs at the fetlock joints. The hairs around the small ergots are now over two inches in length.
The chestnuts on the fore legs in the Zebra are large and smooth, and on a level with the skin; in Romulus and Remus they are also large, and hardly if at all above the level of the skin, but they occasionally give off thin scales. In Brenda the front chestnuts, though relatively nearly as large as in a Zebra, project as far above the level of the skin as in a pure Clydesdale foal. The left hind leg carries a small prominent chestnut about a quarter of an inch in diameter, but there is no rudiment of a chestnut on the right hind leg. The hoofs are the hoofs of a Zebra, and considerably smaller than would be the hoofs of a Clydesdale foal of the same age. They are wide behind and rounded in front, but the bars are relatively short, i.e. they do not extend as far back as the frog. I may add, the nostrils are in their shape a little less Zebra-like than in the other hybrids; that the muzzle suggests the dam more than the sire, the lower lip being, as in the dam, somewhat long; and that the rounded ears are tipped with white, as is occasionally the case in dun ponies as well as in Zebras. As might have been expected, the trunk and hind quarters are more massive than in Remus, while the shoulders are less upright, and perhaps as a consequence of this the action at all times is less Zebra-like than in any of the other hybrids. As fig. 2 (PI. II.) indicates, there is a "swirl" nearly three inches in length extending down the centre of the face between the eyes. The same figure also indicates fairly well the extent of the marking at the end of the second month. The brow arches (hardly visible in the figure) are nearly as pointed as the frontal arches in a Norwegian pony in my possession, and as in the Amsterdam Quagga. This is very remarkable, as in all the other hybrids the brow stripes form rounded arches. The cervical, and in fact all the other stripes as far as they go, agree with the corresponding stripes of Romulus. In the region of the shoulder the markings are very faint, and over the hind quarters only a few indistinct spots and portions of bands can be detected. The lower parts of the legs are only faintly striped, and even the bars across the forearm and the hock are more obscure than usual. But although none of the stripes are very pronounced, there are, strange to say, faint lines between several of the cervical and vertical body-stripes. These lines suggest "shadow" stripes, and seem to correspond to some of the numerous indistinct vertical stripes seen in Zebra-Ass hybrids. In having faint intermediate vertical stripes, this, on the whole, Horse-like hybrid may be said to be, in at least one respect, more primitive (to have reverted further) than either of the other hybrids already described. If this hybrid continues to thrive, she ought to grow into a powerful, active, shapely cob, about fourteen hands in height, hardier and with more staying power than an ordinary mule.
The Hybrid "Norna."
The most attractive of last summer's crop of hybrids has for its dam a good-looking 11-hands Shetland pony ("Nora"). This pony, which will be six years old in the spring, had a foal in 1895 to a small black prize Shetland pony ("Wallace"). Nora is in many ways a small edition of Mulatto, and her foal Norna may be said to be a small edition of Romulus. When a few days old Norna, in her colouring, movements, and make, was more fascinating than Romulus at a similar age; and now that she has increased from thirty inches (her height when foaled on June 8th) to nearly forty-one inches she looks (notwithstanding her single hoofs) as if she belonged to some bygone age. Norna has been from the first more intelligent than any of her contemporaries, and always very much on the alert without being at all nervous or frightened. She followed her dam through a crowd of some thousands of people on Jubilee Day without any hesitation, or evincing any signs of fear, and she now leads quietly and allows herself to be measured without offering any resistance. At birth Norna generally resembled Romulus, both in colouring, markings, and shape; but her head was relatively smaller, and the ears relatively shorter. There was, however, a very important and interesting difference between Norna and the other hybrids. As already pointed out, the croup and rump of Romulus were at the outset marked by numerous rows of spots having on the whole a transverse direction. When his new coat was completed, in August last, I noticed that many of the spots had united to form somewhat zigzag bands that in their direction agreed closely with the stripes on the hind quarters of the Somali Zebra. In Norna, instead of spots over the hind quarters, there were from the first numerous narrow and hardly at all wavy stripes, which line for line almost agreed with the markings in the Somali Zebra. But, further, many of these all but transverse stripes reached, or all but reached, a stripe running obliquely across the hind quarters in almost the same position as the oblique stripe in the Somali Zebra which I have elsewhere referred to as the upper femoral stripe. The remarkable difference between the markings over the hind quarters of Norna and her sire Matopo, and the equally remarkable resemblance between these markings in Norna and the Somali Zebra, seem to me to throw a flood of light on the relationships of the stripes in the various species and varieties of Zebras, and at the same time strongly to support the view already advanced, that the difference between the stripes of the sire and his various hybrid offspring is in all probability due to atavism or reversion. [ See the 'Veterinarian,' December, 1897] If this is the correct explanation, it follows as a matter of course that at least in the markings the Somali is the most primitive of all the known recent Zebras.
That the hybrids have reverted in at least their markings towards a somewhat remote ancestor: - it may be a common ancestor of both the Horses and Zebras: - is also indicated by the presence of faint "shadow" stripes on the neck. From Matopo having twelve cervical stripes and some Zebras having in addition nine or ten "shadow" stripes, and from Romulus having twice as many stripes as Matopo, it may be inferred the typical number of cervical stripes in Zebras is twenty-four or thereabout. But in Norna, in addition to the twenty-four cervical stripes, there were at least five faint "shadow" stripes. In Zebra-Ass hybrids there are usually many indistinct stripes on the neck and body, and numerous spots over the hind quarters. I consider Zebra-Ass hybrids more primitive in their markings than Zebra-Horse hybrids. In having numerous cervical stripes Norna approaches Zebra-Ass hybrids, and the only explanation of this that occurs to me is that in Norna we have, in the striping of the neck, a further reversion than in any of the other hybrid offspring of Matopo.
During the first three months the mane of Norna was quite upright, though thicker than in the other hybrids. During the last four months the mane has been increasing in length, and it is now no longer upright; the posterior half hangs over to the right side, the part between and in front of the ears forms a thick forelock, while the intermediate portion hangs to the left side.
Norna with her short head, peculiarly tattooed face, and the heavy mane hanging partly to one side and partly to the other, looks very quaint, and seems to differ quite as much from her sire as she does from her dam the black Shetland pony. The coat is now very heavy, the long hairs over the body measuring over three inches, while many of the hairs over the brow are nearly two inches in length. If Norna develops after the fashion of Romulus, she will: - a year hence: - be a compact small striped pony from 11 to 11.2 hands in height. As is the case with Romulus, there is nothing about Norna that suggests either an ordinary mule or a hinny. She has excellent well-formed feet, only a few short hairs at the fetlock, and not a rudiment of warts on the hind legs.
The Hybrid "Heckla."
Heckla's dam is a 12-hands skewbald Iceland pony. There is so much white in this pony (Tundra) and the yellow is so pale that I thought her hybrid foal would be nearly as light as a pure-bred Zebra. As it happens, Heckla is the darkest of all the hybrids, and the stripes are nearly as obscure as in the "Clydesdale" hybrid Brenda. As she lay by her dam shortly after birth, she looked like an overgrown Hare with an unusually long head and relatively long ears. From the first her coat has consisted of long coarse hairs, and the warts on the front legs are prominent, as in her dam. Measuring 32½ in. at birth, she was 43 in. at six months, and is now (January 12th) 43½ in.; the circumference of the knee being 9½ in., and the fore-shank 5½ in. Though Heckla has always carried a heavy coat, and is dark in colour with white tips to her ears, she generally agrees with Romulus in her build and markings; but her action is freer, and more like that of a hackney than a Zebra. She promises to be quite as large and as active as Romulus, and more able than Romulus to withstand cold and to flourish under adverse circumstances.
The length of the head and the shortness of the neck suggest that the Iceland ponies belong to a different race than the black Oriental-looking West Highland ponies. They may be direct descendents of the Horses hunted by the men of the Reindeer Period. Their ancestors may have gradually worked their way northwards with the Tundra fauna which then as now lived near the edge of the ice. If Heckla owes her dark colour to reversion, it may be inferred her ancestors were of a mouse-dun colour.
It is too soon to offer any opinion as to whether Romulus or any of the Zebra-mare hybrids will prove fertile or specially useful either at home or abroad, and it is equally impossible to say whether they will withstand the African Tsetse fly, or have better constitutions than either ordinary mules or Asses, but this much may be said, they all seem very hardy. Romulus has been in perfect health from the first, as indeed has been his Zebra sire, while nearly all my mares and Horses have had colds and other ailments. Quite recently the four hybrid foals and three ordinary foals have been suffering from the presence of Strongylus armata. One of the pure-bred foals (Mulatto's second foal to an Arab Horse) died from the effects of the parasite on the 1st of January, and a thoroughbred foal has been reduced almost to a skeleton; but the four young hybrids, though no longer so bright or in so good condition, are evidently rapidly recovering, and will, I trust, be soon all right again.
The editor of the 'Scottish Farmer' believes Romulus "will be invaluable for driving or riding on account of his hardiness," and he has stated that all the hybrids "have feet and legs like whalebone, with the kind of pasterns that Clydesdale men fancy." ['Scottish Farmer,' Nov. 27, 1897] It is well known that Captain Lugard and Major von Wissmann have advocated steps being taken to breed Zebra hybrids.
Captain Lugard, in his work on 'Our East African Empire,' writes:: - "Some years ago I advocated experiments on taming the Zebra, and I especially suggested that an attempt should be made to obtain Zebra mules by Horse or Donkey mares. Such mules I believe would be found excessively hardy and impervious to the 'fly' and to climatic diseases.... I would even go further and say that their export might prove one of the sources of wealth and revenue in the future; for, as every one knows, the paucity of mules both for mountain batteries and for transport purposes has long been one of the gravest difficulties in our otherwise almost perfect Indian Army Corps." Since this was written much information has been gained as to the dreaded Tsetse fly, but apparently there is extremely little chance of Horses being made immune, being so treated by innoculation or otherwise that they will be able to survive if once infected by the peculiar minute organism so intimately associated with the all too fatal disease.
Further, owing to the destruction of cattle by the rinderpest, the transport difficulties have been increased in Africa, while the Frontier wars have enormously increased the demand for mules in India. On the other hand, it has been proved that it is a comparatively simple matter to cross various breeds of mares with a Burchell Zebra, and if experts are to be trusted the hybrids (Zebra-mules as some call them) promise to be as useful and hardy as they are shapely and attractive. The preliminary difficulties having been overcome, it remains for those in authority to take such steps as may be necessary to ascertain of what special use, if any, Zebra hybrids may be in the various parts of the Empire, but more especially in Africa and India.
As I am anxious to obtain as much information as possible bearing on equine hybrids: - on crosses between Zebras, Horses and Asses: - and as to the fertility of the various kinds of hybrids (mules, hinnies, &c), I shall be most grateful for accounts of any experiments hitherto made, more especially with Burchell and other kinds of Zebra. I have not yet heard of ordinary mares having been crossed with Burchell's Zebra in South Africa; but doubtless some of the readers of 'The Zoologist' may be able to give me information on this subject.
|
BACK TO HYBRIDS & MUTANTS INDEX INDEX |