THE PAWPAD MARKINGS AND RELATED ITEMS IN THE SYSTEMATICS AND ECOLOGY OF GENUS FELIS.
Treatises of the Bavarian Academy of Sciences, Mathematical and Natural Science Department New Series, Issue 52, 1943
From the zoological collection of the Bavarian State: The Pawpad Markings and Related Items on the Systematics and Ecology of Genus Felis, by F. Schwangart, with 11 illustrations.
Presented by E. v. Stromer on October 24, 1942.
Munich 1943, Publishing house of the Bavarian Academy of Sciences
On commission by the C. H. Beck'schen Publishing House, Munich
Printed by C. H. Beck’s printing house in Nördlingen.
In my first work dealing with the tribal history and systematics of genus Felis (1928), which was expanded into a brochure in the following year (1929, 1), I already tried to justify my conviction of a closer relationship of groups previously considered to be independent, a northern group, F. silvestris Schreb., the "wild cat" par excellence, a southern one, F. ocreata Gm., "fawn wild cat", and an eastern one, F. ornata Gray. In a monograph about house cats regarding breed formation and descent (1932) I carried out this validation more precisely and specifically included the eastern group in particular. The result was "a geocircle which, with a considerable number of geotypes [landraces], occupies an enormous territory distributed over the three continents of the Old World along with a number of their islands".
[This requires some notes on terminology to begin with. The genus Felis is narrowly defined here, contrary to a frequent and notably older usage. The three groups and closest relatives just mentioned remain in Felis. The "reed cat" (the so-called "swamp cat") Chaus chaus (Güld.), which already stands out from the others, is eliminated; The affiliation of E. LÖNNBERG’s Poliailurus (Art pallidus Büchner) and Eremaelurus thinobius Ognev (1927) remains questionable, and both will be discussed here.
We should agree on the spelling, the mixed Latin-Greek "-ailurus", or the completely Latinized "-aelurus" in Felid names. The authors contradict each other, e.g. Lynch ailurus, but Eremaelurus, or even Poliailurus in LÖNNBERG, but Poliaelurus in OGNEV. In such cases I will remain with the first person who described it.
P. RODE and R. DIDIER (1935) demand BRISBON (Le rčgne animal, therein "sylvestris") 1756 as the author of the name silvestris in place of SCHREBER 1777. Since BRISSON's work appeared before Linnaeus’s Ed. X, it stays with SCHREBER and "silvestris". According to the "Journal of F. Mammalogy" No. 10 (1935) the "German Inst for Mammalian Science" accepted an application from H. POHLE for the definition of scientific domestic animal names for referral to the International Nomenclature Commission. The house cat should then be called F. catus L., not F. domestica. In terms of nomenclature, catus (Linnaeus 1758) has preference over domestica (GMELIN 1788). Owners need not fear difficulties from this change since they use "catus" more often anyway; the designation also permitted the term "catology" for serious cat research, a name for which has long been desired.
According to TH. HALTENORDT (1940), p. 3 note 1, all Felis falling under the term "fawn wild cat" should not be called ocreata Gm but constantina Förster (1780) according to RI POCOCK (Mamm. Of India, vol. 1, London 1939). This change of name created confusion among domestic cat breeders; they have just had to switch from the local form known as maniculata to ocreata. It has often been suggested to refrain from strictly adhering to the priority principle in favour of a final determination of naturalized names in cases where the priority principle is threatened. In response to F. HEIKERTINGER's (Entomol. Vol. 27, 1940; Zool. Anz. Vol. 136, 1941) renewed approval of this procedure, a questionnaire was first asked within the "Journal for Applied Entomology ". Incidentally, HALTENÖRDT himself (1c, p6) took a similar point of view when deciding between silvestris and "ferus" in favour of the former.
The HEIKERTINGER suggestion has the merit of having highlighted a frequent, insufficiently appreciated embarrassment of many zoological practices. If these circles are against the path of "suspension" provided for in the nomenclature rules, which RUD RICHTER reminds us of (in "Continuity of zoological nomenclature - against the rules or with them?", Zool. Num. 139, 1942) against HEIKERTINGER, then because the personalities whose vote decides such cases plead to avoid applied science and its needs. One may judge whether this mistrust is right or wrong after the commencement of my proceedings, first of all to keep the naturalized name ocreata instead of using constantina until the planned decision. I also advocate the appropriate participation of the relevant discipline, which is competent on a case-by-case basis, in the committees that are entrusted with such questions.]
Some of the feasible distinguishing features between silvestris and ocreata did not hold up at all then, and most of the rest only suited the extremely well-appointed forms far from the areas of contact or approach. Several of these features recur here; a compilation of them can be found in the works mentioned.
Regarding my scepticism about fundamental distinguishing features, especially between F. silvestris and ocreata, I had a predecessor in R. I. Pocock to a degree. In his work "On English Domestic Cats" (1907), which was also valuable for the cat breeding, he believed that the following external differences between the two wild groups could still be recognized in their originally assumed scope, which I did not take into account in my previous publications and they are otherwise hardly known: "In F. sylvestris the four paired stripes on the head and neck are well defined ; on the occipital region they diverge from the middle line and run backwards almost to the shoulder as four wavy widely separated stripes, the ad median pair being, as a rule at all events, better emphasised than the laterals, which lie quite at the sides of the upper surface of the neck. In F. ocreata the head- and neck-stripes are usually badly defined; when present on the neck they are narrow and lie close together."
What he adds on both sides of the pelt drawing is already marked as transitional by his own text. According to my present material, the same applies to the features mentioned in the above quotation. In later publications by Pocock I do not find any comments on his attempt at separation. Our more recent research has accepted my position that silvestris, ocreata and ornata are closely related, represented e.g. by O. KLEINSCHMIBT (1938) and TH. HALTORDT (1940/41), this when he says: "One has to agree with Schwangart (1932) when he advocates taking the concept of the actual wildcat very broadly and not only the northern, central and southern European representatives of the Felis silvestris group, but also those of most of the Mediterranean islands, the forms of the African black cat group. . . and that of the Asiatic F. ornata with their attributable subspecies." Similar to P. Rohe and R. Didier (1935), without naming myself, Haltenordt adds another reference to this grouping, which is based on the microscopic hair structure, whose structure M. P. Rode (1931) and the same with Didier (1935) used it specifically for the Felid systematics. Among other things, two species of the same genus can be distinguished, Profelis aurata (Temm.) and temminckii (Vig. and Horsf.), the close association of two supposedly independent species, namely F. silvestris and ocreata, raise an objection to the obvious conclusion that the old world Felis and the South American lynchailurus are closely related, a question that will be touched upon here.
In my work on South American felids (194t) I drew attention to other possibilities of these systematics, including the importance of the penis structure, especially its tip doming. That this is not functionally necessary was shown to me by the case of a breeding tomcat (wild-coloured tabby "Milo"), who remained fully capable of breeding after surgical removal of the glans tip. Furthermore, I would like to suggest paying attention to the structure of the first two cervical vertebrae, which according to the results of Stromer von Reichenbach (1902) are of value for the Carnivore systematics.
Haltenordt (1 c) cannot agree with my view (1932), according to which some of the features (hair, tail, ears, extremities, structure) stand out in contrasts in the geo-circle Felis are due to climate conditions can be assigned to Bergmann's and Allen's rules. He criticizes the "completely inadequate material". However, I believe I can refer back to my reasoning for this view as well and also draw attention to data from this work, including the parallel case of the South American lynchailurus that will be described.
If the genus Felis is considered as a geocircle (so-called "geographical racial group"), a question arises regarding the naming of the individual forms. If one puts the geocircle on the level of a "species", of course without prejudice to its genus rank, one must choose the oldest of the previous species names as a unifying species name; that would be silvestris, and we would then speak of Felis silvestris silvestris, Felis silvestris ocreata, ornata etc. Partial types that were subordinate to one of these groups would be treated in a quaternary manner: F. silvestris ocreata maniculata etc. I’ll leave it here with the style F. silvestris such as grampia, F. ocreata such as maniculata etc.
The preparatory work for the following results was based partly on material from our collection, and partly on my travels as a judge at international cat shows abroad, where I took the opportunity to acquaint myself with museum material. Since there were only a few days available at the individual locations, with only a few hours available in between strenuous judging work, I asked for particularly instructive pieces from the collections to be worked on in Munich and compared them with related material from our own and other German collections. As far as there were South American cats, an initial outcome of my analysis is given in my treatise on South American cats, etc. (1941), which is to be continued. As far as the Felis genus is concerned, some of the material was used in my work on the Manul (1936). Another particularly valuable contribution was made by shipments and insightful comments by Prince Altrecht of Bavaria, from the time when he was busy setting up the Yugoslav hunting industry. If the circumstances of the war had not compelled him to end this activity, even more significant advantages would have resulted for my work and other work at our museum, with which the aforementioned had long been connected by his scientific interest. We would like to express our special thanks to him! I have already given thanks to the foreign museums, with the exception of one case of further valuable additions that should also be mentioned here, that of Rhenish, Spanish, Caucasian and Algerian material from the Alexander König Museum in Bonn, whose director, Dr. von Jordans, unselfishly helped me in the current times of difficult transportation.
Next to the matter of the markings of the sole as one of the most common criteria for differentiating between the Felis species, several other such characteristics arise here, as already mentioned in the title. It is useful to anticipate two of these at this point: the question of the fertility of mongrel animals between typical, or regarded as typical, members of the three Felis sectors, or between those and domestic cats and the colour of the nose leather. The last objections that could be raised against a continuous transition between representatives of the three sectors are based on these two.
Mongrels of European wildcats of Silvestris type with African wildcats as well as with domestic cats have often been bred. As an example of the first case, R. J. Pococks hybrid of F. silvestris (male) with F. ocreata ugandae Schwann (female) (Pocock, 1907, 2), as an example of the second B. Peters' mongrel (1932) of Bulgarian F. silvestris with domestic cats can be mentioned. Peters has analysed the inheritance of traits insofar as it is possible on the basis of first-generation offspring. I reproduced Pocock's illustration of his hybrids in my work from 1928/29.
The comparative results on the distinguishing features of the Felis representatives in the Silvestris, Ocreata and also in the Ornata sector had already invalidated the doubts about the fertility of hybrid animals to such an extent that experimental proof was actually only missing for the sake of methodical "elegance". The "Research Centre for German Wildlife" (Fritz Schmidt, 1942) has now provided this evidence for F. silvestris X domestic cats. I wrote about this in the same place (1942). Harz region breeding material has been used for wild cats. We can say with confidence the passage mentioned that it was pure-blood. The great similarity of one of the pieces held there with some apparently Silvestris-blood domestic cats can be shown by a comparison of two pictures, one of which (Fig. 1) is taken from the essay by F. Schmidt, while the other (Fig. 2) is mine from 1938 and comes from the same magazine. The striking similarity between the two animals is based on details of its shape and pattern; the fact that this very large house cat is domestic, as shown by its yellow-red ground colour is irrelevant here; there are cats of this variety in every colour, including those in wild colours; the shape, pattern and ground colour all together give my "wild-coloured grey tiger-striped", see illustrations in my monograph from 1932. In any case, I can advise repeating the very simple breeding experiment using wild material from an area that has a greater amount of typical species than the Harz. The Eifel would be particularly suitable, where, according to Haltenordt (1940, 1), the already rich and secure population has increased since the ban on leghold traps, whereas in the Harz "its existence is well assured, but nothing is known about its frequency". The essay by F. Schmidt includes pictures, all of which only show wild cats. Science and cat-breeding would have a great interest in pictures of descendants of the hybrid animals, as a more precise description of these remains to be drawn up.
Contrary to the lack of concern in some hunting areas, it should be noted that a definitive judgment as to whether an animal is a house cat, hybrid or wild cat, can be impossible, and not only on material from regions with smaller wildcat breeds, such as Mediterranean countries, but also here with fully grown animals of either type. Just consider that F. silvestris has mongrelised with the old shorthair cats from the south (Nehring, Pocock, F. Schwangart) and that there is now a large proportion of silvestris even in longhair domestic cats (F. Schwangart). It would be easy to confuse these with powerful semi-angoras, which are quite common in the country, and pictures such as those of "Silvester" and "Simsoh" in my monograph from 1932 show how domestic cats can reach wildcat size. In addition, there are certainly frequent hybrid cats and there are also domestic cats that have been outside for long periods of time and have grown a thicker fur as a result.
According to the investigations by Kirk (1935) and some supplementary cases by Haltenordt (1940), the old statement of a longer small intestine in domestic cats is confirmed as a plausible consequence of the household diet. For this purpose, tests on larger sets of material are desirable. Material is needed for the various breeds of house cat, especially the longhaired ones, and very young animals, in order to clarify the probable inheritance of these acquired changes. Material is needed for wildcats from changed environments and climates. In practical terms, it is advisable to measure the intestines of all dubious specimens killed in the wild and to report the result, along with the external measurements and with remarks about the appearance, to the scientific authorities, if one is unable to send them the intact specimen.
From what has just been said, one can see without further ado that wildcats, especially when not fully grown, are just as easily mistaken for house cats as other way round, one more reason for hunters who have been instructed to be hostile to cats should be less trigger-happy against the "pest".
For African felis of the typical Ocreata type, the ability to produce fertile hybrid offspring with domestic cats is natural since these form the main strain of domestic cats.
Gray (1867) described Indian house cats which he considered hybrids of pure (ocreata-blood) house cats with Chaus chaus (Güdenst), the "lynx cat", to which he assigned his "Chaus ornatus" due to similarities such as the possession of ear tufts and the relatively short tail. The former characteristic also occurs in other Felis, especially African wildcats and also among our house cats. The second characteristic distinguishes the typical silvestris. Thereafter, there will have been hybrids with Felis ornata or purely ornata-blood domestic cats. Ognev (1930) mentions the case of a mixed breed of domestic cat and F. ornata. I am not aware of any attempts to deliberately breed such hybrids.
As the once commonly used criteria to distinguish between F. silvestris and ocreata lost their value a characteristic which was mostly set aside became more significant: the colour of the nose leather - "red nose" for silvestris, black for ocreata. Ornata remained undiscussed. The fact that it is precisely this feature that has been overlooked in collating works of the last few years (Ognev 1930, Pocock 1934) and in detailed descriptions such as Eckstein (1919), in which the colouring of the bridge of the nose is often precisely noted, is certainly due to the fact that the authors were not aware of any live animals. E. Schwarz (1930) mentions the "very red nose" among the few characteristics that made him consider describing his Balearic cat F. catus jordansi as a "real wildcat", while his two specimens were otherwise "of the African type" and F. ocreata mauritana (Cabrera) appeared to him to be closely related, circumstances that led him to consider this Balearic cat a bridge between the northern and southern Felis. But contrary to his belief, the "red nose," as well as the black nose, occurs in domestic cats, especially in those with a grey pattern – with both the ancestral striped pattern and the domesticated marble pattern - the wild nature of this interesting cat remains uncertain although likely, as I'll show later. As a result, however, it will also emerge that more and more of the supposed special forms of the Mediterranean countries, described from only a few specimens, are equally suspect, as Pocock (1934) has already shown for the case of F. agrius Bäte from Crete.
In my monograph (1932) I gave preliminary statistics on the colouring of the nose leather in domestic cats and tried to relate this peculiarity to their changing colour characteristics. In them, the deep red reaches "an intensity that is not surpassed in wild cats". This red colour occurs in all degrees, right down to pale pink in albinism. Regarding the origin of this colour, Silvestris blood and domestic albinism both come under consideration. The occurrence of cats with red-black spotting on the nose surface should be added, as this red always seems to be albinotic in nature. Regarding its role in breeding and other details relating specifically to the domestic cat, I can refer back to the work mentioned.
Among the few living representatives of European and African Felis and the few prepared ones where it was still discernible, all specimens from the area usually assigned to Silvestris had the red nose leather, all Africans, plus the island form F. sarda Lataste, had black nose leather. Contrary to other characteristics, however, this characteristic does not decide an animal’s specific classification.
The texts of the descriptions of forms of the eastern group Felis ornata (Gray) also fail with regard to this characteristic, e.g. because Ögnev (1930) differentiates them from silvestris. Gray's illustration (1874, Fig. No. VI) of his "Chaus caudatus" (= Felis ornata caudata), which belongs to this region, shows the "red nose", but also a fully solid sole strip as is usually the case with African types. If in L. Zukowsky's description of his Felis caudata longipilis (1914), according to the name change by the author (1921) and according to a more recent classification to F. ornata macrothrix Zuk., it says "the nose is brownish in colour, not yellowish as in F. caudata" (type specimen), this means the bridge of the nose, which, by the way, usually corresponds to the "red nose" in domestic cats. So when asked, Mr Zukowsky wrote to me that, as he remembered, the nose leather of his two ornata forms described in 1914, macrothrix and griseoflava, whose representatives he saw alive, were red. Representatives of the ornata sector in the Felis circle seem to mediate between the image of typical silvestris and typical ocreata, according to the nose leather and the colour of the sole.
[Of the forms that adjoin the silvestris-ocreata-ornata geocircle to the east and partially overlap with ornata, Poliailurus Lörmberg (1 c) and Eremaelurus Ögnev (1 e) are considered to be closely related to it, perhaps also belonging to Felis. Chaus is already out, the Prionailurus are more distant, the Manul (Otocolobus) belongs to the lynx in my opinion (SCHWANGART 1936). - LÖNNBEHG bases his generic and species description of Poliailurus pallidus (E. BÜCHNERS Felis p. 1894) on the skull only, for the external characteristics he refers to the describer of the species. He does not mention the nose leather. Likewise, the two describers of Eremaelurus thinobius, OGNEV (1930) and BILKEVIC (1934) ignore it.
With regard to Eremaelurus, which OGNEV put between Felis and the Manul, I have already voiced doubts in my work on the Manul (1936), as mentioned here. The position between two so distant genera, which is evident from the first description, was improbable. The mainstay of its closeness to the Manul was the supposedly rounded shape of the pupil in the of light, in contrast to the slit-like narrowing of the Felis pupil. Now, according to his work written in Russian, whose content I translated myself, S. BILKEVIC (1934) found that living specimens of this trans-Caspian "Sand cat" contracted their pupils into a slit, just like Felis. Even after everything else, it can be assumed this cat can be assigned to Felis.]
But focusing now on my original theme, the markings of the sole, I will first turn to the southern Felis sector i.e. the "ocreatas".
I found two statements in the literature about a lack of the typical sole strip of the hind legs. Gray (1867) writes in the British Museum about a series of skins from the North African "booted lynx", F. caligata, which was considered a distinct species at that time and which has since been affiliated with ocreata, most of them have the back of the hind feet black, but it is "not a consistent characteristic", for several specimens were paler at the back (the dorsal stripes), and in some of these the blackening or browning of the sole was limited to a "point on the outer corner". It can also be seen from this that this criterion was discussed before A. Nehring (1888), who is considered the first who wanted to distinguish between wild and domestic cats (1888). R. I. Pocock (1907) spoke out against the universal validity of this feature, mentioning that not only some domestic cats, but also some "Egyptian cats" lack the sole-stripe.
I myself examined eight skins of long-legged and slender, very large desert cats from the Cyrenaica in the Turin Museum - Benghazi, Derra, Uadi? - the private collection Festa and found the otherwise strongly prominent sole-stripes as well as dorsal stripes on the reddish-tinted specimens. A specimen belonging to the museum called "Colonia Erythrea", also tall, long-legged, slim, large, in all a typical ocreata, had the sole-stripe interspersed with light spots.
A collection of nine Sardinian cats (F. sarda Lataste, usually understood to be ocreata) from the Milan museum, two of which came into our possession through exchange, show the sole-stripe throughout; in some it is interspersed with slightly lighter areas. The same is true of a Berlin series that I mentioned in 1928/29. In Turin I saw the following Sardinian cats: No. 1550 and 1588 of the Sassari Museum, a gift from Sign. Enzo Bizzozero 1891, both of them conspicuously large compared to the average of the mostly small Sardinians, almost identical in appearance, with a strong reddish base colour and stripes that stand out clearly, as well as the particularly long, black end zone of the tail; with a brightly indented, therefore incomplete sole-stripe. Furthermore, an unnumbered piece by Arzana, pale, a little reddish, with only an indistinct sole-stripe. [In my work from 1929 I named the Sardinians as an example of flavism or erythrism that had already begun in the wild cat. At that time, I determined its "occurrence" in the above-mentioned series of Berlin skins. This is not a consistent characteristic of F. ocreata sarda Lataste, there are also extremely grey backgrounded ones.]
In the Paris Museum d’Histoire Naturelle I saw specimen of Felis silvestris reyi Lavauden 1929, from the Biguglia lagoon near Bastia (Corsica). It's a relatively small animal. The hair and tail are similar to the Sardinians. A sole-stripe is present. The specimen is very dark and mottled on a grey background, with a conspicuously long, black end zone on the tail. The ears have small end tufts, as is usually the case with Africans, seldom with F. silvestris, usually with Sardinians, quite often with house cats. Regarded as a wildcat, the sarda specimen, which it most closely resembles, is placed between the Silvestris type and the Ocreata type, despite lagging behind the previously measured Sardinians in terms of tail length, thus Haltenördt (1940) decided it belonged as a silvestris subspecies. In addition, it is female; according to Haltenordt's information about German wildcats, the females among the Mediterranean ones are likely to have shorter tails on average. Of 10 Sardinian skins, which differ quite a lot in tail length (Milan Museum and ours), the reyi type comes closest to one with full length to tail length ration = 95:32, against 83:27 for reyi (according to Lavauden). Incidentally, some of these Sardinians are even smaller than the reyi type, also addressed here. According to the markings, this is a suspected domestic cat. The weak skull structure suggests either this or the Sardinian type. The rarity of Corsican material in comparison with Sardinian material is remarkable. I would like to remind you of my communication in the work from 1928/29 about a wild cat with the appearance of an African ocreata in the museum in Nice, which is known there as the "Riviera cat". The mounting of that stuffed specimen prevented an examination of the rear sole.
The mounting of this stuffed piece is instructive to me in that a complete sole-stripe is present in the very powerfulg specimen of an Ocreata male from the Abyssinian highlands (pelt No. 1532, skull No. 2639 of the Civico Museo in Milan, location Azozo, 2160 m above sea level, gift from U. Ignesti), whose overall appearance must have resembled that of a continental European Silvestris, also in terms of size, strength, especially the teeth, zygomatic arch spacing, etc. the skull, is almost equal to the largest of our German wildcat skulls. I wrote down the following about the fur: cream-grey mottled, centre of back, centre of head, muzzle broadly reddish-brown, just faint horizontal stripes radiating from that zone down into the belly hair. This creamy-grey, shaggy. Upper side short, straw-like, sides with woolly texture. Tail grey, somewhat woolly, long-haired, moderately long, in the terminal half with black, rings around it and moderately long, black tip, appearing rather thin despite its shaggy. Hind thighs more strongly streaked. Complete, deep brown sole-stripe. Ears moderately large; brown-red above, with only a suggestion of ear tufts (in the form of an extended row of hairs at the edge, as with most F. silvestris). Length 118, tail 39 cm. Described by the local museum: Felis ocreata maniculata Temm. In terms of fur character, colour details, proportions, and skulls, there are strong echoes of our silvestris. On the other hand, the tail shape, the (albeit moderate) size of the ears, especially the complete sole-stripe, testify to it an African ocreata. The traits suggestive of our wildcat are certainly climatic. The determination as a sub-form maniculata probably cannot apply. The establishment of a new form and its systematic evaluation is dependent on further material, and this would have to be compared with sufficient material from other African highlands, e.g. from the Atlas mountains. - Among those now available to me, three skulls and a skin from the Alexander König Museum in Bonn come under consideration, labelled P. Spatz collection, Smendon near Constantine, Algeria, and surely Felis catus mauretanicus Cabr. The fur (male), which will also be mentioned here in another context, has echoes of the typical silvestris, such as the somewhat woolly hair character, and especially the ears which are too small on the Ocreata scale. The tail is short-haired, thin and long, the sole-stripe of the hind feet is complete. The two adult male skulls are clearly inferior to the Abyssinian in resemblance to those of typical silvestris.
On the whole, the Ocreata group, plus the Sardinians, in terms of its sole stripes in the previous term, despite transitional individual variants. The Silvestris group is different with regard to the sole patch, right into the zones of the continental pattern shape.
I am dealing here first with the areas neighbouring the ocreata range, the European Mediterranean countries. After their sole-markings, the Sardinians already join the Africans with regard to the rule and the variants. For the Corsica and the Riviera cats this question remains open.
From my on-the-spot notes relating to Italy and its neighbourhood, here is what is relevant to the matter.
1. Gift of ornithologist G. Martorelli, in the Milan Museum, additional type specimen of his Maremma cat (F. maniculata var. Mediterranea, from Populonia on the coast) and according to him identical to the Sardinian cat. The piece is a "slate-colour tiger" in the sense of my breeding system of domestic cats, with a prominent pattern, and in my opinion a domestic cat. - M. Hilzheimer (1913) says of Martorelli "Nota Zoologica sopra i gatti selvatici e loro affinita colle razze domestice [Zoological note on wild cats and their affinity with domestic breeds]", Milan 1896 (without further details) "Martorelli has . . advocated a polyphyletic origin of European domestic cats. From the Tuscan Maremma and Sardinia he described a F. mediterranea close to F. maniculata and wanted to see to see part of the domestic cat's descent in it."
2. Pocock (1934) does not mention this case, but mentions F. molisana, Altobello Abruzzo, Molise, Central Italy (Fauna dell’Abruzzo, Mamm. P. 35, 1921, and Monitore Zool. Ital., Rediconto Mamm. P. 35). Pocock considers the existence of this breed between F. silvestris silvestris in Tuscany and the same in southern Italy to be unlikely, without considering the altitude possibilities. Th. Haltenordt (1940) presents them as F. s. silvestris. - In the Turin museum I saw a prepared hide without a skull from the Abruzzo, Festa collection, about which I noted: yellow-grey, wild-coloured, blurred stripes, rows of spots, more prominent than in the Sardinians. Light throat, whitish from lower lip downwards. Plain stripe on the back in the rear half. Belly cream coloured, a whitish tinge in the region of the belly spots. Tail not bushier than body hair, which is quite long and somewhat woolly. The sole-marking is somewhat elongated. Back length about 54, tail length strong 26 cm. The specimen has a lot more of silvestris than the Sardinians but is not a typical F. silvestris. Domestic cat mongrel cannot be ruled out.
3. Milan Museum, "F. silvestris Schreb., Sibari (Calabria), presented by Restelli". - Stuffed, very similar to the Sardinians, closer to ocreata than to silvestris. As with the Sardinians, the tail is somewhat bushier than that of a typical ocreata. Sole-spot is large, elongated. Domestic cat or mongrel probably excluded. The piece thus appears as an intermediate between silvestris / ocreata.
4. Milan Museum, "Gatto selvatico dintorni da Zara, Inverno 1920, R. Perlim vend., Cranio manca [Wild cat from Zadar region, Winter 1920, R. Perlim vend., Skull missing]". - Large animal, prominently striped lengthways on the back. Fur plush. Northern thick, short, broadly ringed tail with a very blunt tuft. Mottled, slightly paler towards the top. Sole-stripe. Ear brush suggested. Stripes almost absent with the exception of the dorsal stripe. Overall impression, apart from the sole-stripe, is that of a strong, typical silvestris.
5. Milan Museum, "Colle di Tenda, Gatto selvatico, No. II. II cranio manca. M. Giulino ded. [Tenda hills, Wildcat, Nr. II. The skull is missing. M. Giulino dcd.]". - Fur dark, tawny, basal part of the hair grey. Stripes blackish. Around the throat there are two strong, dark half-rings. Ears red, with blackish tufts in them. There are no chest and belly spots (as is the case with a minority of German F. silvestris). Strong, short silvestris tail. Continuous, complete, strongly contrasting sole-stripe. Whole length 91, tail with tuft 34 cm. - A second hide (No. I) from the same place is a typical silvestris and is very different in colour from the one just described, as is another stuffed specimen from the museum. - The one described here could, despite its dimensions and predominantly silvestris characteristics, be a mongrel with the domestic cat.
6. Civico Museo in Turin, No. 1715. Province of Cuneo. Typical silvestris. Pale, back stripes almost disappeared, sole-stripe inwardly indicated. Presumably alpine (Scottish or Ligurian Alps).
7. Turin Museum, Pelt No. 1678, Skull No. 4973, Gatto selvatico [wild cat], Regio Mandria (near Turin); low hills, c. 300 m above sea level. Strongly striped, straw-like fur, tail much thinner than in the previous Turin specimen. Sole-stripe prominent, but narrow and lightly interspersed. Relatively small animal; in my opinion, not a domestic cat.
8. Turin Museum, No. 1574. "Bosco dell’Incoronata, preso Foggia, Apulia. Ded. Prof. Griffini 1899." - From a maximum of 50 m above sea level. Quite a strong animal, size like our silvestris from Transylvania. Fur character predominantly woolly and straw-like. Throat with a pure white crescent. No belly marking. Basic colour predominantly grey, rump stripes pale but visible. A partially interrupted and split dorsal stripe. Tail relatively short, ending slightly bushy. A distinct sole-stripe. - Despite its origin from the extreme south of Italy and its sole-stripes, the animal has Silvestris characteristics throughout. I certainly think it's a wildcat.
The evidence given here probably proves the urgency of methodically working through of the entire Felis material in Italian collections and, in addition, a methodical investigation of the Felid fauna of the Mediterranean continental and island world. I made this demand in my publications of 1928/29 (1928, p. 21). The samples I have just given are also far from being sufficient for establishing individual forms, which one may be tempted to do just like predecessors in this field. The temptation to rush into systematics could easily be reduced by taking better account of those descriptions with which new names are not already linked. Such communications are generally neglected, as the literature clearly shows. - The same variability as in the sole-markings shows this small amount of material with regard to other characteristics used to distinguish between silvestris and ocreata, such as to establish sub-types. Even if, for example, samples from the southern Alpine regions (Tenda hills, Kottische or Ligurian Alps) form contrasts with those from the neighbouring lowlands, whereby the typical alpine form seems "northern", looking at others among my samples teaches us that even a purely altitude justification of all details is not enough.
As an appendix to the Italian Felis types, attention should be drawn to the results of excavations by F. Zorzi (1940) near the northern beach of Lake Garda. These are late Neolithic pile dwellings. Among the human and animal remains, those from Canis familiaris intermedius Woldrich, Canis familiaris cfr palustris Rütimeyer and, according to the author's determination, from Felis silvestris Schreb, were found as a rarity. Zorzi refers to my inspection of the two cat skull remains and to my opinion expressed here that it is either a Mediterranean wild form closer to the Ocreata type or a domestic cat of that time. The proportions and the strength of the bone fragments presented to me permit only these two possibilities.
The material sent to me by Director Dr A von Jordans gave me an insight into the Spanish area of the F. silvestris in the common concept ascribed to the sector. Miller (1912) claims to have identified a specifically Spanish F. silvestris tartessia in addition to F. silvestris silvestris, the first of which was known from northern Spain, the second from south of the Ebro, from central and southern Spain. Pocock (1934) and Th. Haltenordt (1940) have included this classification in their lists of F. silvestris. Cabrera (Fauna iberica, 1914) gave a picture of tartessia, according to Haltenordt. I was unable to complete this work. The F. catus jordansi, Balearic cat described by Schwarz (1930), also belongs here as Spanish.
The Bonn skins and pelt, some with skulls, are geographically divided into two groups: 4 of them come from Lagumilla near Bejar in the hilly valley between Sierra de Gata and Sierra de Gredo, south of Salamanca (i.e. north-western central Spain), at an altitude of probably around 1000 m. - 2 pieces from Morqueruela near Teruel, in eastern Spain and a little further south, occupies an even higher position in the southern part of the Iberian foothills. I also received two unworked skulls from Bonn from the area around Linares de Riofrio in the foothills of the Guadarama, Province of Salamanca. These come into consideration regarding the question of "tartessia".
Regarding the first group:
Most robust sample, adult male, February 9, 1934. Overall length 93 cm, tail 31 cm. With a silvery background colour that characterizes these pieces, as does the entire colour and marking type in general. The pattern is quite striking, especially the stripes on the head and back. The tail is thick and bushy like a Silvestris, but the hind feet have an almost complete sole-stripe. The skull is silvestris-like, with relatively small zygomatic width. – Specimen by specimen, the aforementioned silvery tinge in the background colour and decreasing body size are more and more pronounced. Classification of all samples: F. catus silvestris Brisson.
2. adult male, the accompanying skull dated – 18.9.1934. Dimensions according to the original label: "Body 52 cm, tail 26 cm, ear 7 in, sole 11 cm." At 10 cm, the sole-stripe can be considered complete. In this and the following samples, it is interspersed with a bit of silver in the upper part, corresponding to the overall background colour. The skull similar to the previous one.
3. female, 28.10.1933. Original note; "Body 52 cm, tail 28 cm, ear 7.5 cm, sole 11 cm." The sole stripe with 8-9 cm can be considered complete. It is consistently dark in this specimen. Skull missing.
4. female, 7.10.1934. Original label: "Body 47 cm, tail 23 cm, ear 7 cm, sole 10 cm." Sole-stripe is 8-9 cm so complete. Its upper part is interspersed with silver. Juvenile skull.
The fur character of these 4 pieces when shot at different times of the year is consistently long and soft. Throat and belly spot present in all, but individually different sizes.
To the second group of mainland Spanish samples mentioned, from Morquerela near Teruel:
1. female, 14.2.1930. The attached original dimensions: "Body 48 cm, tail 24 cm, ear 4 cm, sole 10.5 cm." Overall, it is similar to those from the Bojar area. Dorsal stripes a little closer set, hair shorter, not so soft, more straw-like. A pale sole-stripe is present, but interspersed with light areas, with a uniform lower area that can be understood to be a sole-spot. White spot on the throat, not on the belly, whitish tint on the chin. Tail strawy. Skull still juvenile.
2. adult? male, 14.12.1930? Total length about 52 cm, tail 28 cm. Here the horizontal stripes are much closer together. Hair like the previous one on the torso and tail. Marking of the hind sole is a spot with an interrupted smoky extension in the direction of the heel. Throat and belly spots are missing. In both samples there are strongly pronounced reddish tones, especially on the underside, more so in this one. Skull missing.
Both groups, from the area of Bojar in western Spain and Teruel in eastern central Spain, both from mountainous terrain, are intermediate between "silvestris" and "ocreata" types, although these are more characteristic of the western ones, with regard to the sole-marking but approximate the eastern representatives. It is possible to speak of a sub-form of silvestris for both groups.
The type area of the F. silvestris tartessia described by Miller (1907) is Goto Donana near Jerez de la Frontera. In Pocock’s report (1934) there is another sample from "Madrid". The descriptions of the small amount of material cannot convince me that this form is independent, especially in view of the uncommon modifiability of all the characteristics of Felis: it is larger than F. s. silvestris, darker in colour, has longer teeth, and the sagittal crest of the skull extends from the coronal suture to the occiput, which is otherwise not found in silvestris. P. Rode and Didier (1935) have now described a silvestris sample from the area of Vesoul (Haute-Saone, France) that is as large as the "Tartessia" specimen - I saw the sample, labelled silvestris, in the Paris museum - and even among our scant German material there is a powerful male specimen from the Black Forest (Krieg Collection), the sagittal crest of which is the same as that of the Haute-Saone specimen. The author, Miller, himself explains that "tartessia" is "difficult or impossible to distinguish" from Scottish wildcat ("F. silvestris grampia") . — Of the two Spanish unprepared skulls from Bonn mentioned, one is still subadult, the adult lags behind our mostv robust German specimens in size, which is matched by the attached fur measurements: "Body 57 cm, tail 21 cm, ear 6 cm, sole 9 cm." Both these and the other Spanish ones described above are similar in conformation. They do not have a continuous sagittal crest. Certainly, the material in the Bonn museum can be considered more correctly as a special sub-form, perhaps it also contains two of these, which geographically intervene naturally between the European silvestris type and the African ocreata type, similar to the Sardinians in this regard, but with a greater resemblance to silvestris.
If one includes the "tartessia" specimens, one arrives at a peculiar distribution of Felis in Spain, from which fact, just as for the Italian area, there arises an urgent need for better research and for inspection of the entire museum stock:
1. The western European silvestris protrudes into northern Spain, up to about the Ebro (London material).
2. The Bojar-Teruel line would have a smaller home range with transitional features towards ocreata, especially from the sole-marking to the complete sole-stripe; this line presumably continues, as I shall show, to the Balearic Islands, with changes tending further towards ocreata.
3. In southernmost Spain, there are again typical silvestris (so-called "tartessia") represented by large and strong specimens, which, according to Pocock, should also fit into the line given for the second group (Madrid skull).
Of the two examples of the Felis catus jordansi E. Schwarz 1930 (Bonn Museum) from Mallorca that I have, one is the type specimen, the other must be considered an additional type specimen according to the author's statement, according to which both skins "matched in every detail".
This second specimen is labelled "San Juan, Mallorca, male, leg. Parpal, 9.6.1930, Paratype". The skull is missing. The fur that I have, whose support was about the size of the type, is poorly preserved; the hairs thinned, in places depilated. Much more light-yellow tint than the type, tail long and thin. Hind foot with a somewhat elongated marking on the sole. Most likely a yellowish-grey, dull tabby domestic cat.
The type is described as such; withfurther information: "Sta. Margerita, Mallorca, male, leg. Parpal, 13.1.1929 ". The total size of the pelt of about 75 cm (nose to tail tip inclusive) is that of a smaller house cat, the skull justifies the author's designation as subadult. His designation as male contradicts that as female on the label; I consider the latter to be correct. (Exact determination uncertain.) Moreover, if the hair is a winter coat it cannot be considered "very long" on a wildcat scale, as the author intends. It lies within the range of my "wild-coloured short-haired" domestic cat. I have already spoken about the "very red nose" as not being a characteristic of domestic cats. "Intense coloured ears" are also common enough for them. For example, in addition to a golden-yellow front edge and a light red-brown bridge of the nose to the "wild red" of the nose leather, a beautiful grey striped tabby that I keep (1942) resembles the jordansi type in all of these characteristics. The fact that "the domestic cat of the Balearic Islands" was also present does not matter when it comes to the question of wild or domestic cats, and in those areas feral domestic cats and hybrid cats would be frequent enough. The large ears in comparison with the findings on the Spanish specimens I examined may indicate domestic cats as well as ocreata. All the colouring and markings also occurs in domestic cats. By the way, the "strongly striking leg markings" hardly surpasses the Spanish mainland wild cat specimens.
Taken together, these features are still important for assessment: an incomplete sole-stripe slightly interspersed with lighter hair, the absence of throat and belly spot (with a silvery whitish chin), the bright reddish brown tone of the entire undercoat, which is directly visible on the underside, the very narrow transverse stripes that are blurred in the mottling of the hair, the relatively thin tail. The author himself does not include the skull dimensions in the diagnosis. The zygomatic arch width is small.
Although a domestic cat cannot be ruled out for this interesting specimen, as is so often the case, and therefore more material is necessary, I tend to designate it as a wild cat on the basis of comparison with material from the Spanish mainland and an Algerian skin from the Bonn material that I have. E. Schwarz himself pointed out the relationship between the Balcaron cat and F. ocreata mauritana. Because of this bridging position of the Mediterranean wildcats between the silvestris type and the ocreata type, he would like to concede two names anyway and to have all sub-forms on both sides designated as species. He speaks of F. catus mauritana, F. catus jordansi, F. catus sarda, etc. (The name catus, as already mentioned, is given to domestic cats.)
To briefly touch on this question here: forms such as sarda and, if it is wildcat, jordansi are actually neither ocreata nor silvestris, and closer knowledge will multiply such forms. This point of view is already included in my view of a common silvestris-ocreata-ornata geocircle, which presupposes the existence of bridging types. Miller (as early as 1907) rightly referred to the Sardinians as "Felis sarda", not as a subspecies of ocreata, which it is otherwise considered to be, nor of silvestris, under which E. Schwarz (1 c) places it. On the other hand, there are also forms, such as most or all Africans and continental Europeans, which one must summarize more closely despite the invalidity of many of the previously valid characteristics and leave them as lower categories of ocreata or silvestris. If more abundant material confirms the character I suspected of the mainland Spanish specimens just described as special form(s), I would perhaps assign them to silvestris, despite their tendency to a sole-stripe, for example. The ornata group seems to me to be the closest to complete resolution.
Among the Spanish samples, jordansi is most similar to the piece from Merqueruela near Teruel, which I cited as No. 2. The two pieces correspond in most of the details so that one must regard them as borderline types of a form if they came from a single place of discovery. The only difference is the ear size: the continental specimen has the small ears of the silvestris, the island one has the large ears of the ocreatas and most domestic cats. The two samples from Merquerela differ much more conspicuously in terms of differences in the distance between the stripes, the colour of the undercoat, the absence or presence of a throat patch, one of the many examples of how such small details in individual specimens justify the creation of new forms. In any case, the pieces of evidence from Spain form a clearly recognizable line from mid-west to mid-east Spain to Mallorca (apart from details such as the particularly pronounced sole-stripe in western Spanish samples) – considering the wild nature of jordansi.
If one examines the aforementioned Algerian specimen, this line seems to continue to northwest Africa. It is labelled: F. catus mauretanicus Cabr. Adult male. Coll. A. Spatz, 12.3.1913, Smendon near Constantine, Algeria. - Total length of this fully grown fur 94-93 cm, tail length 36 cm. The transverse stripes have almost disappeared in the mottling of the rather woolly hair. The background colour is very similar to that of the jordansi type and to the aforementioned specimen from Merqueruela, also in terms of the intense reddish brown of the lower hair. The tail, of the same hair type, is somewhat thinner and more pointed than that one. The unbroken sole stripe is unanimously black-brown. No throat or belly mark. The ears are silvestris-like small, much smaller than those of jordansi. The skull shows zygomatic arches, which project in front, with a relatively narrow snout and joint capsule width, as in many Africans and fewer Europeans, and also contrasted with the Spanish and jordansi specimens I visited. A second, adult, also male skull from the same museum, from the same place of discovery, shows these differences even more significantly. A third, juvenile, had an even more infantile, wider muzzle.
When compared with jordansi and the last-mentioned mainland Spanish sample, the overall result is an advance towards the ocreata type. Only the small ear size stands in the way of this. This as well as the slightly woolly texture of the hair is related to the home of its bearer in the Atlas Mountains. In the prominence of the zygomatic arches, these skulls surpass the one described on p. 10 from the Abyssinian mountainous region, which on this point, at least secondarily, comes closer to the silvestris form.
I could not form an opinion about the Greek and Asian continental cats and the Greek island cats. It is unlikely that silvestris in terms of the pattern type has spread as far as the archipelago. However, previous evidence of supposedly special forms has not been proven: the independence of Felis catus morea Reichenbach is disputed (see Haltenordt, 1940), F. agrius from Crete is a domestic cat according to Pocock.
An expansion of the black sole to form a sole-stripe can also be found in the central areas of the Silvestris sector. When K. Eckstein (1919) says of the German wildcat that the "sole of the hind legs is either rusty brown-grey", in which "the colour at the base of the fifth toe darkens to a short, sometimes blurred, sometimes sharply circumscribed spot" or it is "all black", this already includes the phases up to a stripe. The variations are even more clearly expressed in the author's detailed description of the hind foot on p. 49. The frequently asymmetrical darkening he mentions also reveals variations. His associated statement that the domestic cat, "regardless of its overall colour, has a black sole in its entire extent", is based on an error that is hard to understand. Pocock (1934) knows of a wildcat from Salavon near Nancy that "has most of the sole sooty"; according to its overall appearance, he places the specimen in relation to Miller's (1912) Central and Western European "race" F. silvestris silvestris Schreb. The pieces from the Tenda hills that I have already mentioned in connection with the Mediterranean wildcats also belong, by their sole-markings, here.
I received a valuable collection of Rhenish wild cat skins from the Bonn museum. These skins are also instructive with regard to the modifiability of the wildcat markings. Details of the marking patterns should not be considered here; evaluating them requires a larger sample of material from the entire European range of silvestris. For this reason, the skulls are not included here either. The fur characteristics, the length and shape of the tail, the hair and the size and shape of the ears (small narrow ears) are all typical of silvestris. Here:
1. "Felis catus L., Sinzig a. Rh, male, 7.6.1917". Typical restriction of sole-marking. Throat and very faint spot on the abdomen.
2. Also. "30.6.1916". Almost complete sole-stripe. Very large throat spot, chin very bright, weak belly spot. Striking markings, but certainly a wild cat.
3. "Felis catus L., male, label Fleischmann, Antweiler, Rhine province, 27.10.1913." Sole-marking as in No. 1, larger belly spot.
4. "Felis catus silvestris Schreber, male, label. Forester Budde, Pfaffenwiesbach, Taunus, winter 1898". Throat patch spreading over the chin, belly patch. Sole-marking like the previous one.
5. "Felis catus silvestris Schreber. Label H. H. Böker, 22.12.1935. Wehr, Mayen district, Eifel." Sole-marking as previous. Tiny throat spot, indistinct belly spot.
6. "Felis silvestris Schreber, female, leg. A. Schoenberg, Mayschoss, 10.9.1937. Killed in the rabbit hutch." - The sole-marking is slightly extended upwards; the sole is smoky. Throat and belly spot. The basic shade is silvery. Pattern very dark and distinctive. Slender specimen. May be domestic cat hybrid.
Concerningthe following mention of Balkarian and Carpathian material, the distinction made by Haltenordt (1940) between a western Felis silvestris silvestris Schreber and an eastern F. silvestris ferox Martorelli 1896 should be remembered. O. Kleinschmidt (1938) made a contribution to the delineation between the two on the basis of examining West and Central German material, including some from the Bonn museum. According to this, specimens from the Eifel and Moselle region belong to the first type, those from the Harz and Thuringia region to the second. Haltenordt states the "probable" area of ??the first, according to the literature, as: "Northern Spain, France, Western Germany (Rhineland regions), southwest Germany, Switzerland and Italy ", as for the second he stated:" Germany from the Harz and Thuringia region eastwards, General Governorate for the Occupied Polish Region, Russia, Southeast Germany and the Balkan countries." He also places the aforementioned F. catus morea Reichenbach here, but without any guarantee.
A decision on the question of this separation cannot be made here. What I know of supposedly differentiating characteristics is subject to the considerable modifiability of the genus Felis on both sides. Kleinschmidt found an unequal upper and lower limit on both sides by comparing the extremes and using the "carpet method" on the basis of the intensity of the basic colour and the distance between the stripes and based the separation of forms on this finding. Often enough, however, one finds similar differences when comparing any series - examples can already be found within the entire material and area I have named here - and only very large series from a wide area could create foundations that do not, when applying these procedures which are justified in themselves, to these and other individually modifiable characteristics, lead to illusions and contradictions between different criteria. While an excellent breeding biologist like Antonius (in his handbook 1922) has pointed out the "explosive" variability of shapes in dogs, which is already hinted at in its wild ancestors, the same applies to colouring and markings in domestic cats - a circumstance that increase the value of their, in many ways unique, position for science - and this tendency for these parts to change is already a moderate peculiarity of their wild paths. The systematist must also keep this in mind. - For the purposes of this work it is sufficient to combine the assumed forms silvestris silvestris and silvestris ferox under the common basic form of the "typical Silvestris type" and to disregard the form "ferox".
In a row of eleven silvestris specimens from our state collection from Transylvania, the black-brown of the rear sole varies from a narrowly delimited spot as in Eckstein to an extent far up the sole (in two pieces), whereby it is then somewhat paler and slightly interspersed with lighter hair. You can already see it as a broad stripe. Everything else in the overall impression is typical silvestris.
In March 1940 we received from Prince Albrecht of Bavaria two furs from wild cats from Petrovcic near Surcin in the Belgrade area (1940, no. 28, male, and 1940, no. 126, female), both adult. Descriptions:
1. Male. Shaggy hair, lion yellow, slightly light grey mottled background colour; multiple narrow stripes on the head; poor dorsal markings, several pale and interrupted stripes in the front third, a narrow, blackish longitudinal stripe in the rear third, which merges onto the tail, clearly detached. Rear three ringlets and the tassel of the short tail blackish, two of them dark yellow, only suggested. Ends of the front soles slightly sooty, sole-markings on the hind feet. (The ears are missing.) Total length 102, of which tail (with tassel) 31 cm.
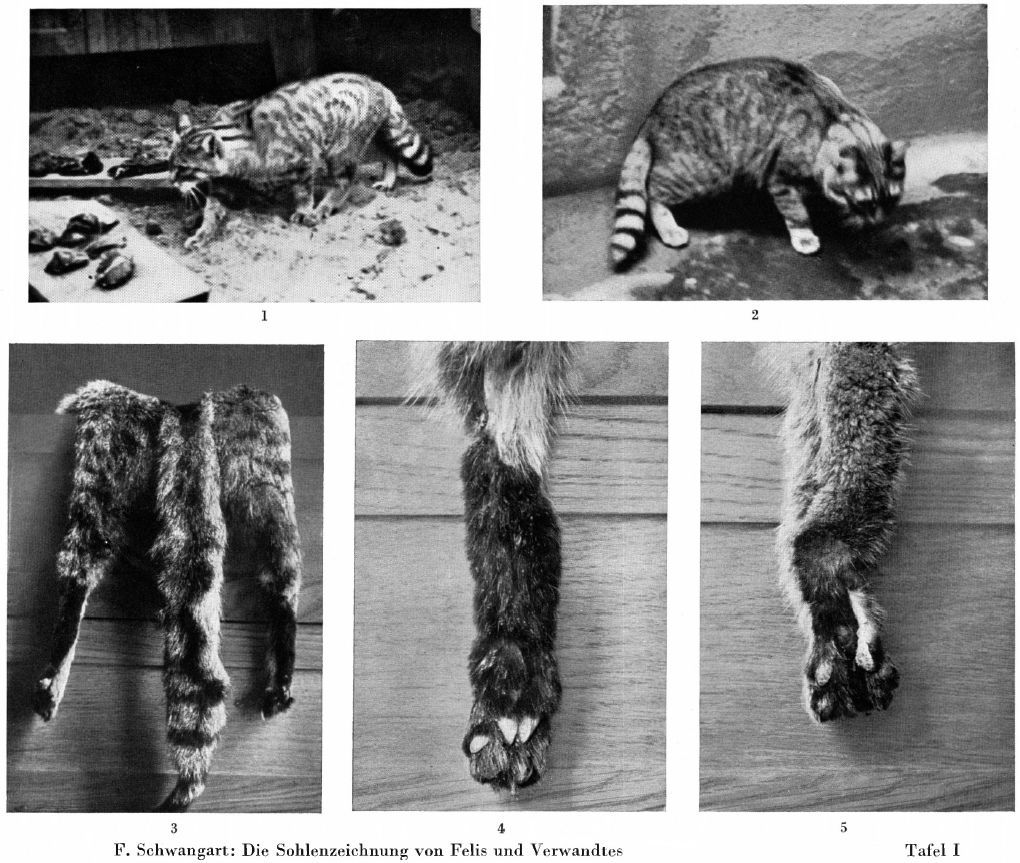
2. Female. Hair a little longer. Basic colour a little darker. The dorsal stripe is wider, faded, does not extend over the tail. Transverse stripes are missing. The rings of the short tail are largely indistinct, split, limited to the upper middle part, representing 5-6 rings. Blackening of the tail tassel longer than the previous one. Forefeet with terminal spot. Hind feet with deep, black-brown, strongly contrasting, almost complete sole-stripe upwards. Ears very small. Whole length 104 cm, of which tail 32 cm. - In spite of the sole-stripes, there is no doubt about the nature of this second sample as a pure-blooded wildcat.
For comparison, my Fig. 3 shows the two hind legs and tail of F sarda Lataste, male, from Tortoli, Sardinia, Zoolog. Munich State Collection, 1935, No. 67. The hair is strawy, in contrast to the majority of African forms, for example the Libyan desert form, and similar to that of the piece I mentioned from Upper Abyssinia; if you consider Central and Northern European silvestris, it rests between their norm and that of those desert cats. The tail is long and thin, almost like theirs, the legs are medium-slender, the hind feet have a sharply defined, complete sole-stripe. - Fig. 4 shows the hind foot of the female sample just described (No. 2 of the descriptions) from the Belgrade area. The long shaggy hair of the typical silvestris is visible at the top of the picture, underneath there is a sharply contrasting, deep black-brown, almost complete sole-stripe, in contrast to the sole-patch of the normal silvestris. - Fig. 5, rear foot of the male sample from the Belgrade area described under 1; slightly damaged towards the bottom, which was limited to the distal end of the sole, but somewhat elongated sole-marking clearly recognizable.
Regarding the pattern of the forefeet, which was not taken into account in the systemics, it should generally be said that a striking dark tone on the underside corresponds in its extension to that of the hindfeet, even if there is hardly ever a complete longitudinal stripe. Among the three samples especially listed here, a brown marking extends furthest upwards in the Sardinian example, in the Belgrade male the marking is limited to the lowest part, in the Belgrade female with sole-stripes on the hind feet it goes a little further so that the pattern of the forefoot behaves as "intermediate", in contrast to the ocreata-like marking of her hind feet. On the matter of the sole-pattern on the hind feet of the two Belgrade samples he had sent, the collector, Prince Albrecht, wrote a letter after he left the office there. His collection of wild cat hides and unprepared skeletons, which was intended for us and was laid out in Belgrade, had been destroyed by the events of the war, as was a living specimen that had been brought intact for my "position for cat research" with the German Society for Animal Psychology. But the collector was able to explain with certainty: "All the pieces that I saw from the corner in front of the confluence between the Sava and Danube rivers were of the same type as the two specimens", i.e. the Belgrade samples I just described. "They all had an elongated sole-patch up to a regular sole-stripe. I saw two example from Pancevo, that is, very close, only the other side of the Danube. These had a small, black silvestris-type markings on the sole. The relatively large number of live wild cats that I had seen in the Danube-Drau corner had, as far as I could observe in the open air, a sole-marking rather than a stripe. So it seems that this approach to the ocreata type is either a local form in the Danube-Sava corner or perhaps it is spreads even further towards Croatia and Serbia. It would be interesting to get more samples from these areas.
A welcome addition to Prince Albrecht's findings on Serbian wildcats was the receipt of two Bulgarian and one Romanian sample from the Bonn museum. Findings:
1. "F. silvestris silvestris Schreber, female, from H. Wolf, Kara Bonar, Winter 1937/38, Strandja-Balkan, Bulgaria." – Silvestris form. Throat spot, belly spot. Very small marking on the sole.
2. Likewise. Very similar to the previous one. Sole-marking a little wider.
3. "F. silvestris silvestris Brisson, male, adult Gek. von Fritsche, 10.1933, Fagaras, Romania."
A very powerful animal. Pattern not very pronounced. Typical sole-marking. Throat patch consists of only a few hairs, belly patch of average sizes. - The thick but not very large skull has a surprising appearance due to its relative shortness, which is related to a strong bulge of the zygomatic arches reminiscent of many African samples.
None of these three Balkan pieces had anything approaching a stripe on the rear sole.
According to what the two Belgrade pieces reveal to us in connection with the collector's explanation, variants with more developed sole-stripes seem to be increasing in certain areas with more limited populations, as apparently is the case in the corner of the Danube-Sava confluence, while in neighbouring areas the sole-patch remains dominant. Perhaps the Spanish samples in the Bonn museum already described belong here. Regarding the collector's assumption that the tendency to a sole-stripe may persist even further into Croatia and Serbia, I refer to the above-mentioned piece with sole-stripe from Zara, but without wishing to conclude anything. According to the collector, the size of the sole-patch in the region where it alone occurs should be smaller than the patch on the sample Nol described here from the variant zone. Strong rivers would be involved in the demarcation between two forms.
Among the eastern forms of the Felis genus, OGNEV (1930) keeps F. Silvestris caucasica Satunin (1905) with silvestris, and its distribution extends from the Caucasus to Turkestan and into Asia Minor; typical locality Borjom (Transcaucasia), as can be deduced from the text of Satunin, although an exact designation is missing from there. The conformation is based on features of the skull. Several authors (Pocock, Rode and Didier, Schwangart) have expressed their opposition to this type of basis within Felis in general and even more so within a sector of Felis, in this case within silvestris. Pocock (1934) thinks that it is "impossible to determine characteristics on the basis of the skull to distinguish between wild and domestic cats", and that he "only found quantitative differences" between that of silvestris and sarda. Felis is an "osteologically extremely modifiable form". Based on my own experience, I can subscribe to his judgment. Most of the describers, however, have tried most of all with craniological efforts on materials from a few specimens. It is the same with the other feline genera I am familiar with, and differences in skulls from genus to genus can also be questionable. Examples of this are given in my work on South American small cats (1941). The usability of characteristics derived from subtle skull anatomy in big cats in Haltenordt (1937) to clarify the relationship under higher systematic categories is under discussion.
Ognev adds a brief description of the coat for caucasica, without taking into account the sole-markings and, as already mentioned, the colour of the "nose". He himself comes to the judgment that certain of the Satunin samples are obsolete on the basis of a comparison of three(!)other silvestris examples with Caucasian fur characteristics, so that in this regard he finds "no stable differences". Pocock draws attention to the mottled rows of the Caucasians he knows, but Ognev's description already shows that it is "about transverse stripes which" only "sometimes turn into a number of spots", as is often the case with our wild cats and even with domestic cats. The silvestris-like Felis of the east, where they border on the pure spotted ornatas, require closer investigation.
I too can only contribute inadequately. In my house cat monograph (1932) I made a few comments, in my 1928/29 pamphlet I noticed the absence of a white belly spot on a number of Caucasians in the Berlin Museum, with the exception of one piece that had a poorly developed belly spot. At that time, I interpreted this as one of the signs of the inconstancy of the "silvestis marks" (white patch on the throat and on the belly), for which there was new evidence. Ognev (1 c) even states that "very white spots" are "not uncommon" also "on the points". Among the material sent to me by the Bonn Museum is a piece "Felis catus caucasicus Sat, female, labelled Th. Pleske, Psif, Kuban region (Caucasus), 10.10.1912". The marking pattern is very much inferior with this one, more than with any of the Balkan pieces. The tail is very long by silvestris standards. There is a pronounced throat patch and a fainter abdominal patch. There is a small patch on the sole. (The paws are missing.) The adult skull is relatively narrow, as is the width of the zygomatic arch.
The regular replacement of the stripes with distinct spots is a main feature of the ornate group Felis, which represents the main eastern part. Pocock (1934) suspects an Asiatic origin for the entire Felis group, and in favour of this he points out the spotted pattern of the Asians known to him, from which the stripes of the western and southern Felis was derived. He agrees with van Bemmelen's theory of the primacy of blotchy rows in mammal markings (J. F. van B., 1931), but he does not name this predecessor. The dissolution of the striped pattern into spots also occurs in the torso region of other wild Felis, continental and island cats, e.g. the Sardinians, but then only partially, not continuously, clearly aligned in stripes and in the form of large, not small spots, like the pattern displayed in ornatas. The only similar case that occurred to me is the domestic cat class (not "breed") of the "Perlkatzen [stippled, tipped cats]" of my system. Incidentally, I only found these in the shorthair cats and do not believe that I have been deceived by the hazy marking drawing patterns on the torsos of the most similar angoras. We have no information about the coat pattern in the border area between the silvestris and ornata sectors.
The South African Felis nigripes pelt, which R.I. Pocock (1907, 3) says is a case of spotting in the ocreata region: " On the dorsal area the arrangement of the spots is sometimes more or less longitudinally linear; and on the sides of the thorax two or three oblique stripes are sometimes formed." The illustration of a living specimen in the London Zoo, and an even clearer image of a fur, show a sequence almost entirely based on stripes. On the living animal, the pattern appears to be more extensive than on the fur. There is a perfect sole stripe on the hind feet. Pocock is certainly right in his interpretation of this as a dwarf form of ocreata. It is considered the smallest of the living felids. Features of the skull appear juvenile, a trait that persists in this cat as it does in other dwarf forms among the mammals. [In my experience of cats, owners, especially women, often ask for a "dwarf cat." Isolated dwarf specimens of the domestic cat appear and there have been various attempts at breeding such strains, but so far in vain. Felis nigripes is certainly easy to domesticate, it could provide a healthy breeding basis.]
F. nigripes represents, when counted as part of ocreata, the only case of white among Africans to date. In its case, according to Pocock (1 c), "the entire underside is white or whitish". The silvestrises and the ornatas behave in the same way as the white element varies in extent. In the ornata shawiana form (Blanford, 1879), according to Zukowsky (1914), "the underside is white", ornata kozlovi has "a pure white chin and same colour throat", in contrast to Zukowsky's ornata macrothrix and ornata griseoflava, which lack all white, as does the Gray’s ornata ornata and ornata caudata. The absence of white was the exception in the silvestris area; as a rule, it has a white throat and belly spot, exceptionally it spreads to the chin, a large part of the underside and the points. The geographical distribution more and less allows the conclusion that white is more likely to be absent from silvestris as it approaches the ocreata sector.
I have mentioned that Felis ornata caudata (Gray), according to an apparently good picture by its describer, possesses a complete sole-stripe, as is the rule in ocreata. The text clearly confirms this, whereas Gray says nothing about the colour of the sole in his description of ornata ornata (1867). Elliot (1883) quotes the original description for F. shawiana Blanford (1879), which is now added to the ornatas, in which it says: "A deep black patch on the back of the tarsus", but it depicts a sole-stripe; likewise for ornata, caudata and catus-silvestris, which also shows the inaccuracy of some of his illustrations in using the example of that last type. For example, in addition to all of these, F. caffra even had a red "nose". L. Zukowsky (1914) found in his Felis caudata griseoflava and his F. caudata macrothrix (according to the later renaming for F. caudata longipilis) "on the rear surface of the heel joint" or "rear side of the heel" is an "elongated black spot" and adds to his note that this is known from Gray’s typical caudata. I have reproduced his picture of two living macrothrix (1932, fig. No. 9); unfortunately, the sole-marking of these crouching animals is not visible. According to the author's description, it was an elongated marking. The following applies to his other ornata forms: murgabensis has "brown-black rear soles" (1914, II), matschiei "black-coloured soles", maimannah "soles of the front and rear feet brownish-black"; in this last case, since the front feet are also indicated, a sole-marking is also a question. - In any case, according to the information available, the sole-stripe of the hind feet appears to predominate in the ornata sector. This behaviour in the colouring of the soles, whereby these ornatas are assigned to the ocreata type is, as already explained on p. 8, in contrast to the "red nose" of at least some forms, as that represents a silvestris trait.
Differences in hair length among the ornatas are noteworthy. The two specimens of macrothrix Zuk. The two specimens of macrothrix Zuk (reproduced by me in 1932) reproduced in a picture by Zukowsky (1914, 1) are reminiscent of the silvestris type with their pronounced long hair, especially when the bushy tails that are not visible in the picture are taken into account. In terms of hair length, these cats even surpass the Silvestris model. Likewise, matschiei zuk. has an "extremely long-haired coat" and, as a result, indistinct spots. It's a little shorter in griseoflava Zuk. These three are then in clear contrast to the short-haired murgabensis Zuk., shawiana Blanford and ornata Gray. In a letter, Mr. Zukowsky confirmed my view (expressed in 1932) that this, contrary to Ognev’s opinion (1930), which presupposes a seasonal difference, is a permanent contrast, and indeed a purely climatic one, as he had already convincingly demonstrated. The animals shown here differ from the silvestris model in their more delicate physique, more pointed face, somewhat larger ears, which are smaller than the ocreata model, and their small spots. These characteristics, with the exception of the last, are often found in long-haired domestic cats ("Angoras"), which do not meet the breed standards (i.e. those for the "Persian" and my "German Longhair"), and the same applies as for these according to Vosseler (1927) from selected Longhairs in Ankara ("Angora"); well-bred strains also tend to revert to these form more often. I then expressed the assumption that the "Angora" that came to us from Asia (originally Persia?) could have come from an old branch of the ornatas line (1932). Pocock (1934) turns against this contribution by ornata by subscribing to the view that our domestic cats are "two-stemmed" descendants of both ocreata and silvestris. He gives no reasons for rejecting my opinion, which I hereby uphold, and does not name the originator of his imagined notion. I also remind you of information given on p. 7 of this work about mongrels of domestic cats with ornatas.
With regard to the tail lengths, according to Zukowsky's precise information (1914, 2), there are again differences equal to those between the silvestris and the ocreata models. "Maimanah's cat" (his F. ornata maimanah) "just like shawiana has a short tail, F. caudata, F. matschiei and F. murgabensis all have a long tail."
According to the criteria of sole-markings, colour of the nose leather, tail length, hair length and character, tinge of white, and presumably also according to the ear size, we see a divergence in the ornatas compared to the silvestris and the ocreata models - there will be as little lack of transitional forms here as there is between those models - while the character of the marking pattern on the trunk holds the group together to some extent. There are obviously no significant differences between the skulls and the silvestris and ocreata material. In the sense of an approximation of ornata-placed forms to F. silvestris, Zukowsky finds, by comparing his griseoflava and macrothrix against kozlovi Satunin 1904 from the Turpan Basin (East Tienshan), that its overall impression appears closer to silvestris than to, for example, F. ornata caudata.
It would be premature to speculate about the cohesion or contradiction of the various features mentioned here in the individual ornata forms described so far due to uncertain information and insufficient evidence. Overall, however, the assumption can be made that within the ornate group there are contrasts in the range between the silvestris and ocreata models and that a division of the ornata group will be necessary.
Let us add briefly to the distribution of the ornata assigned to Felis: It extends through widely different climates; altitudinally up to significant heights, "up to 2000 m", as Zukowsky states from one of his forms. To the west, adjoining the Russian F silvestris (see Ognev 1 c) and perhaps overlapping with them, "ornata" can be found in Mesopotamia, Persia, Afghanistan, Turkestan (Tarim Basin and further east), southwards to central India. The subspecies or races that have been established are always based on only a few pieces. They are rarities in our museums. Nine are named. Ognev (1 c) finds some of them are identical with one another. He lined up the Zukowsky forms we were just talking about: griseoflava belongs to ornata caudata Gray, murgabensis and matschiei remain independent, macrothrix appears synonymous with the dubious schnietnikowi Birula. Maimanah is not mentioned by him as it is not Russian. The existence of valid objections to the inclusions contained herein has already emerged from my discussion of Zukowsky's forms, but a detailed revision is not the task of this work.
To return briefly to two closely related Eastern forms:
Bilkevic (1 c) describes the front sole of the trans-Caspian "sand cat" Eremaelurus thinobius Ognev, which belongs to the Felis circle or is quite close to it (see here page 9), but not the rear sole. The front sole has a wide, dark zone. A sole-stripe is then to be assumed at the back.
For F. Büchner's steppe cat Felis pallida, which F. Lönnberg (1 c) established as a special subgenus Poliailurus due to skull differences, which appear considerable here, the description of the first author remains authoritative with regard to its appearance. According to this, "the sole-colour is blackish", indicating the formation of a streak. The fur seems to be rather short, the body colour similar to the mottled yellow-grey of African desert cats, Eremaelurus or to the English domestic cat called "Abyssinians". Büchner relates the distinguishing features to Chaus, including the longer but still relatively short, bushy tail on this scale, which is similar to that of silvestris or ornata macrothrix. The specimens of the two researchers came from the central Chinese province of Kangsu; Büchner's cat is referred to as a mountain dweller, from the South Tetung Range.
Domestic cats may lack a sole-stripe; it may be replaced by a sole patch of very different dimensions up to an incomplete stripe, and almost replaced by a completely light-coloured sole in the background-colour. This is due to two different causes: silvestris ancestry or domestic influence. Among the grey is the change between stripes and spots along with intermediate stages on the domestic "classic tabby pattern" ("marbled", "blotched" Pococks) as well as on the basically different, transverse-striped wild pattern. Red and black nose leather in domestic cats can combine with markings and stripes on the sole. I have already dealt with other criteria for differentiating between felid forms, including domestic cats, in this work and in my work of 1932. I will return to this matter at the end.
In an analysis of English domestic cats, Pocock (1907, l) corrects at length a number of errors arising from the lack of knowledge of "cats from the London streets". Examples in this work demonstrate the same deficiencies that can be disastrous to the performance of systematists. As I have proven several times and can corroborate it through further, drastic cases, this deficiency is related to a disdain for the subject, which certain circles, especially our one-sided bird protection sport, has brought into the audience, including the scholarly world. A thorough knowledge of the domestic cat is a prerequisite for successful research into the Felis genus and the Felidae beyond.
In the previously published first part of my systematic monograph on the South American small cats (1941), I mentioned the eastern "Pampas cats" of the Lynchailurus Severtzow genus (1858), which have which have the same name as a subgenus and which are represented by several subspecies of the genus Lynchailurus (Lynch.) pajeros (Desmarest) distinguished as: Lynchailurus (Lynch.) Pajeros pajeros (Destn.) Emend. F. Schwangart, Lynchailurus (Lynch.) Pajeros hyperthrix n. Sp. and Lynchailurus (Lynch.) pajeros braccatus (Cope) emend. F. Schwangart. The latter has three "phases" which differ in the colouration of the feet in a similar way to the representatives of the ocreata and silvestris sectors in the old world genus Felis. Between a phase of broadly streaked, deep blackening of the rear sole (phase C) and one with a darkened patch that only forms a single spot (phase A), there is a middle phase (phase B) with an imperfect blackening that clearly exceeds the condition of A. This intermediate stage has so far only been documented by two skins - one bought in Assuncion (Paraguay) by H. Krieg, the other from the Barbieux collection, probably from Matto Grosso - about whose origin it can only be said that they must come from two far apart territories, whereas those I know of phase A from more southerly regions, those of phase C from the hill country and low mountain range of that area which forms the northernmost and the closest to the equator of the range of the overall type of the pajeros geocircle.
On the basis of the few pieces that document the three phases, however, a decision as to whether these are geographically determined differences cannot yet be made; hence the neutral expression "phases". In any case, the remarkable analogy to the state of Felis has so far emerged in that the blackening of the rear soles is most pronounced in specimens from the warmest climates, i.e. in the southern ocreata group, while in the more temperate climates the blackening may be weakened. The middle phase is represented here by the area where there is perfect blackening. It corresponds to the geographical conditions if the pigmentation increases in one case (Felis) southwards and in the other (Lynchailurus Lynch.) northwards. The ornatas of the eastern group of Felis remain out of scope until more material is available.
The colouring of the hind foot of the three phases in Lynchailurus (Lynch.) pajeros braccatus can be seen in the theoretical work (1941) on the fur pictures no. I reproduce them more clearly from the same subjects here in Figs. 6, 7 and 8. It is also worth mentioning that the soft-furred, partly long-haired forms Lynchailurus (Lynch.) pajeros pajeros (Desm.) emend Schwangart and Lynchailurus (Lynch.) pajeros hyperthrix F. Schwangart 1941, living further south, do not have any blackening of the feet, so in the lack of sole-pigmentation they go beyond the norm of Felis silvestris. There is also a perfect parallel between these South American cats and the old world Felis in many details of the rest of their structure and type i.e. in the decrease in tail-length and ear-size moving from tropical to moderately cold climates. In the case of Felis this line extends northwards, in the case of Lynchailurus (Lynch.) it extends southwards to Tierra del Fuego. So, if further evidence is required for my previously mentioned opinion the fact that these relationships in Felis correspond to Bergmann's or Allen's rule (as shown in my work from 1932) should be included in the parallel between genus Felis and subgenus Lynchailurus
In the case of the Felis, as I have said, the difference in the sole-marking of the hind feet corresponds to that, albeit less extensive, of the front soles. The front feet of the three braccata phases shown here show it more perfectly: the foot of phase A (Fig. 9) has only the usual stripes without longitudinal blackening, the foot of phase B (Fig. 10) has intensified stripes connected by a strong, black longitudinal stripe, the base of phase C (Fig. 11) has a uniform black tint that completely encompasses it.
Text image no. 16 in my work of 1941, a head sketch of a Lynchailurus Lynch. braccatus, as well as some of the panel images of various living animals of the genus, reveal great similarity of their overall appearance with that of the old-world Felis, though they are of other new-world cat species, the "Round-eared" cats and stand out through their cone ears. One is tempted to assume that the Lynchailurus belonged to an independent group of Asian immigrants, namely those of Felis descent, whereas the ocelots and smaller American spotted cats, which are closely related to them, could be descended from the Asian prionailurus of the Bengalensis group which resemble Lynchailurus in many ways. But the assumption of such a close relationship between Felis and Lynchailurus goes against features of their skull conformation which approximates other American cats, as I explained in my work of 1941, and of their hair structure, as M.P. Rode showed when comparing a F. ocreata with Lynchailurus pajeros (Desm.). This phylogenetic question remains open.
MAIN FINDINGS AND CONCLUSIONS
1. Of the common physical criteria used to differentiate broader and narrower subspecies or races of the genus Felis, the following were discussed: the sole colour, the nose leather colour, the ear size, the length and shape of the tail, the distribution of white, the fur character, other differences in the basic colour and pattern, skull shape characteristics. The colouring of the hind sole has been systematically used in the dichotomy of sole-stripe and sole-patch. The first characterizes the silvestris model, the second characterizes the ocreata model. According to my comparisons, the front sole corresponds to a certain extent with the rear. This is evident not only in Felis but also in the South American subgenus Lynchailurus, a side-note regarding sole-markings.
Among the Felis, the Africans, as possessors of sole-stripes, are still somewhat conservative. Exceptions seem to be individual. Even with altitude convergence with our silvestris, the sole-stripe can persist (Upper Abyssinia, Atlas region). The Sardinians follow suit. In the silvestris sector of the Felis geographic circle, not only in the border zone against the ocreatas on the Mediterranean Sea, but also deep in the continent, the sole-pattern changes, through all gradations up to a complete sole-stripe. There seem to be separate local groups with a predominant or general sole-stripe, for example along a line across Spain presumably up to the Balearic Islands - here there is the question of F. jordansi E. Schwarz - and within a region of Yugoslavia. In the area of the eastern sector the stripe seems to predominate, but evidently the extent of the dark area of the sole changes. Grey striped domestic cats and grey marble [classic tabby] domestic cats, both long- and shorthaired, show all levels of sole-marking from stripes through to narrowly delineated spots.
The colour of the nose leather, ignored in systematics, can only rarely be reliably determined on skins. According to some of these and especially the scanty evidence from living animals, it would be black in Africans such as the Sardinians, but red in typical silvestris - this exclusivity is questionable at least for the Mediterranean region - red is at least widespread in the Ornata sector. In no case was this criterion sufficient for distinguishing between F. ocreata and silvestris. F. jordansi (Balearic Islands), if a wild cat, represented the combination of red nose leather and dark sole-stripe among the wild felis. The same probably occurs among ornatas in the eastern sector of Felis. Domestic cats, both longhair and shorthair, have both the black and the red nose leather. A distinction must be made here between the deep wild red colour and the albino-domestic pink colour. There are transitions between the two, and there is also a colour split between red and black. Red and black nose leathers are found in domestic cats in combination with sole-spots and sole-stripes.
The ear size varies throughout, but the Africans have larger ears within the size range, while the continental silvestris has smaller ears. Intermediate examples inhabit the Mediterranean countries and, apparently for altitude reasons, also the mountainous African countries. There is an example of a small-eared specimen from Algeria, and one with medium-sized ears from Upper Abyssinia. Small-eared forms occur in the ornata sector, but this still needs to be clarified. In domestic cats, the ear size fluctuates within wide limits; in the case of longhairs, smaller ears predominate independently of directed breeding.
According to the tail-length, the Africans have the "thin" and "pointed" long tails, and with some variation the Sardinians are, similar. The forms and lengths of tails in those cats assigned to silvestris are uneven, increasing north-south towards longer tail, but with individual variations everywhere, so that there is an overlap with the long-tailed shorthaired domestic cat. The length and shape of the longhaired domestic cat’s tail tend towards the silvestris type. With the ornatas, one can differentiate between long-tailed and short-tailed forms based on images and descriptions (with Zukowsky).
Among the Africans, white was only found in the dwarf form F. nigripes Burschell, where it dominates the entire underside. In the silvestris sector it may also be absent, exceptionally in individual cases, and more and more often to the south where it is probably also typical. The majority of the silvestris have a white spot on the throat and abdomen; there may also be only one of these markings present, or the white may spread to the chin and be more widespread on the underside, including the points. A marked change can also be seen in the ornata sector, from a complete lack of some of the variations described through to a completely white underside. In domestic cats, apart from the widespread leucistic and albino white, the aforementioned two "wild blemishes" often occur.
In terms of hair length and density, the northern and central continental silvestris are far superior to the specimens from African lowlands. In Europe, furriness decreases more and more towards the south, at least at lower elevations, just as in Africa, according to previous evidence, the more densely furred Felis live in higher mountain regions. In neighbouring regions of the Central European Zone, they often show very different fur, but due to the scanty material, a strict verification of altitude or other climatic conditions would have to be carried out on a case by case basis. Our domestic cats show all transitions from a velvet hair similar to that of the African desert cats (comparable to theso-called "Abyssinian" breed in England) and pronounced "angorism", as initiated in some silvestris types and almost perfect in certain long-haired ornate types (e.g. macrothrix Zukowsky), which contrast with the short-haired ornatas from other areas. Contrary to Pocock (1934), I maintain my hypothesis that long-haired ornatas are involved in domestic long-haired cats ("Persian", my "German Longhair").
According to tail length and shape, ear size, fur coat, strength and length of the extremities, a general overview of the genus Felis in its wild forms is subordinated to the Allen and Bergmann rules. I maintain this against the objection of Haltenordt (1940). For more information on the justification, see F. Schwangart (1932). It is possible that factors such as the colour of the nose leather, the extent of white, and the sole-colour also follow these rules. We also have a side-note in the behavior of the South American subgenus Lynchailurus Severtzow. (Schwangart, 1941.) For a more detailed analysis of the environmental impact on all these characteristics, a more detailed investigation of the altitude distribution of forms of the genus Felis is very valuable. This work contains examples of how to approach this. Perhaps the ornata sector would provide the best information on this.
In order to differentiate between more narrowly defined subspecies or "races" of the genus Felis, details of the basic colouration such as the position, density and intensity of the stripes, and differences in the shape of the skull, either on the whole or its parts, have been most widely used, analogous to the procedure for other groups of mammals. The material known up to now probably leads to suspicions about the usability of some such criteria in the justification of narrowly defined forms, furthermore, for example, the contrasting bulkiness of skull structure between the lowland and desert areas corresponding to the traditional African Ocreata form such as the Libyans) against those corresponding to the European Silvestris type - but beyond that, everything else fails. In particular, the popular skull measurements which are often preferred unilaterally, even when justifying a few pieces as novel forms, have the least success. Especially with the Ornata sector, according to available material, the dissolution of tabby stripes into distinct small spots that are not strictly aligned is a characteristic otherwise only found in my domestic "Perlkatzen" [stippled] (so far only short-haired) and which, with large and aligned spots, approaches the African dwarf form F. nigripes. (Despite the impression of a lax alignment, the markings are subject to the same plan as tiger stripes as seen in ornatas and my "Perlkatzen".)
In the generic definition of Felis and other Felids, the study of hair structure has served us well (Rode, 1931, Rode and Didier, 1935). It has confirmed the close association of all these Felis and can serve as an objection in the still open question of their phylogenetic association with the large-eared Lynchailurus of South America. The following must be checked for their value as criteria in the complex of questions about the Felis: the type of penis armament, which in my experience has not proven to be functionally necessary in the domestic cat, the construction of the first two cervical vertebrae (with E. Stromer von Reichenbach, 1902) and that of the hyoid bone.
2. Regarding the systematics of genus (geocircle): I would like to keep to the summary of two extensive silvestris and ocreata groups, sub-forms in their area are then named in a trinary fashion. For example, if the Serbian forms described here, with a predominance of the sole-stripe, turn out to be a closed population. On the other hand, forms that stand out more prominently in their intermediate position due to special features should be given their own "species name", e.g. not F "ocreata sarda", but Felis sarda (probably together with the reyi Lavauden form from Corsica, if these turn out to be purely wild-blood). This systematic evaluation would remain doubtful, e.g. for my Spanish forms in the Bonn material together with jordansi from Mallorca (if purely wild-blooded), with regard to the Africans for types like those described here from Upper Abyssinia (Milan Museum) and Algiers (Bonn). The South African Felis nigripes, dwarf form of ocreata, remains independent. Transitional types such as those discussed above from mainland Italy can, provided there is more material, present us with the same alternative. Whether there is horizontal or vertical [altitude] isolation of a form, it would be treated the same in terms of systematic assessment.
So far one has had the least luck with the list of those far-reaching silvestris types such as the western silvestris silvestris and eastern silvestris ferox Martorelli, the Spanish silvestris tartessia Miller, and the silvestris caucasica Satunin. Using the same differentiation methods, I could have based types on most of the groups, even on individual items, in the descriptions in this work. In this respect, it is desirable that when studying literature, more attention should be paid to descriptions and comments and not immediately to nomenclature.
A total breakdown into equivalent geotypes allows one to recognize the ornata sector, which covers altitudinally and horizontally widely different landscapes and climates, within which there are profound contrasts, even after the scant material available to date. This also includes the decision about Eremaelurus Ognev, which is certainly a Felis and not a link between them and the Manul, and about the closely related but remote subgenus Poliailurus Lönnberg.
3. One of my reasons for describing Italian specimens was to encourage a closer study of Felis in the critical Mediterranean zone and all of the museum material there, as I have tried to do again and again since my work in 1928/29. Research in our immediate vicinity should be supported by contributions as happens with those in remote areas. Only the stroke of luck of being a judge in international cat shows gave me access and insight into these.
Within Italy, including its north, samples often reveal a tendency towards the Ocreata model, but there is also no lack of contrasts, as is the case with Dalmatian and Alpine pieces. - Some details: An original specimen of Martorelli's "Maremma Cat," to which phylogenetic conclusions were attached, is a domestic cat the same as my "slate-blue striped tabby"; F. silvestris reyi Lavauden (the type specimen), which is regarded as a distinct species or subspecies, is a close relative of F. sarda Lataste, if not a hybrid or domestic cat; two cat skulls from late Stone Age pile dwellings north of Lake Garda, which were stated to be silvestris (although the decider, Mr. Zorzi, already pointed to my opinions regarding these), if they belong to the Sardinian type or ocreata-like mainland Italians, could also come from a domestic cat of that time.
4. On the basis of a comparison of shapes and evidence in the literature, on expects to find domestic cat crossbreeding in all three Felis sectors, "silvestris", "ocreata" and "ornata". The full fertility of such hybrids is to be expected; in the German silvestris it has been confirmed by an experiment at the Schorfheide Research Center. However, we recommend a cross-check of wild material from areas with a higher frequency of pure-blooded wild cats, such as the Eifel. There is also still a lack of an exact description and illustration of the mongrel descendants, as well as the determination of inheritance in them.
I have warned here, and also through pictures, against the hunting and scientific optimism with regard to distinguishing between wild cats (all three sectors) and wild-coloured house cats or hybrid cats. Sometimes – as this work gives testimony! - accurate distinction is impossible even for the well-informed. Obvious domestic cats and pure domestic cats have been described by systematics as wild varieties or subspecies. Examples from this work: In Felis jordansi E. Schwarz the secondary type is a domestic cat, the main type is possible; F. maniculata var. Mediterranea, Martorelli's Maremma cat, I consider to be a domestic cat; a piece from Abruzzo, the home of F. molisana Altobello, designated as a wildcat, seems to me to be a hybrid; Felis reyi Lavauden, the type specimen, is suspect at least. Some museum designations of the same type were added. An exact knowledge of all the types of domestic cat and a correct understanding of the scope of its variants is indispensable for the explorer of wild felids, and the same is true beyond the genus Felis! R. I. Pocock has previously provided direct evidence of such knowledge, and many others have proved their ignorance.
5. The wild-patterned domestic cats combine the mostly heritable variability in body shape, hair character, pattern details and colour nuances that occur among wildcats in the Felis geocircle, and domestic cats go beyond those in extremes as well as in details, not to mention the diversity of domestic cat colouring. As regards shape and colour, the same applies to the bearers of the purely domesticated "marble [classic tabby] pattern" of the house cat, which is fundamentally different from the wild striped pattern. The impression is that of an all-round as well as "explosive" "luxurizing", which can only be interpreted as the release of an irrepressible natural urge towards diversity, which was physiologically restricted under the pressure of harsher conditions in the wild and which has now broken through, driven by rules. The considerable modifiability already shows such an drive in the wild material. The systemics must take this into account.
BIBLIOGRAPHY
(Some of the publications mentioned below are labelled in the text)
Antonius, O.: Basic features of a tribal history of domestic animals. G. Fischer publishing house, Jena 1922.
Bemmelen, J. F. van: Colour markings of mammals. In: Verb. D. German. Zool. Ges. Zool. Number 5. Suppl. Leipzig 1931.
Bilkevic, S.: Le Chat deserticole (Eremailurus thinobius Ognew). In: Trav. Inst. Zool. Ac. Sc. del’USSR. Vol. 1. Leningrad 1934.
Blanford, W. T.: (Felis shawiana re.). In: Scientif. Results of the second Jarkend Mission, Mammalia, 1878 (quoted from Ognev, 1930) and: Journ. Asian. Soc. Bengal. Vol. 45, 1876 (quoted from Zukowsky, 1914, 2).
Büchner, E.: About a new cat species (Felis pallida n. Sp.) From China. In: Bull. Ac. Imp. Sc. St. Petersbourg. N. S. Vol. 3 (35). Petersburg 1894.
Eckstein, K.: Contributions to a more detailed knowledge of the wild cat. In: Arch. F. Naturg. 85th year, Dept. A. Berlin 1919.
Elliot, D. G.: A Monograph of the Felidae or Family of the Cats. Taylor and Francis. London 1883.
Gray, J. E.: Notes on certain Species of Cats in the collection of the British Museum. In: Proc. Zool. Soc. London 1867.
Gray, J. E.: On the Step-Cat of Bokhara (Chaus caudatus). In: Proc. Zool. Soc. London 1874.
Haltenordt, Th.: The relationship of the big cats to one another. In: Journal of Mammals, Vol. 12. Berlin 1937.
Haltenordt, Th.: Contributions to the knowledge of the wild cat (Felis silvestris Schreber): systematics, distribution, body dimensions and weight, intestinal length, stomach contents, litter size, growth. In: Headquarters ber. Ges. Naturf. Friends. Berlin 1940.
Haltenordt, Th.: Where do wildcats still live in Germany? ln: The German Hunt, year 1940, Neudamm.
Hilzheimer, M.: Overview of the history of pet research, especially of the last 30 years. II The house cat, In: Annals of Zoology, Vol. 5. Würzburg 1913.
Kirk, J.: Wild and Domestic Cat compared. In: The Scotch Natural. Nr. 216, 1935. (Nach Haltcnordt, 1940, 1.)
Kleinschmidt, O.: The wild cat. In: Forests of the world. P. Parey, Berlin 1937.
Kleinschmidt, O.: The German Wildcats, ln: Falco, born 1934. Halle (Saale).
Lavauden, L.: On the Wild Cat of Corsica. In: Compt. Rend. Ac. Sc. Bd. 189. Paris 1929.
Lönnberg, E.: Notes on some Cats from Eastern Asia with description of a new Subgenus. In: Arkiv för Zool. Bd. 18. Stockholm 1926/27.
Martorelli, G.: Observations on Mammals and Birds made in Sardinia. Frat. Braiolo. Pistoia 1884.
Miller, G. S.: Catalogue of the Mammals of Western Europe. Published by British Museum, London 1912.
Nehring, A.: About domestic and wild cats. In: Humboldt, monthly magazine for total Naturw., Vol. 97. Berlin 1888.
Ogneff, S. J.: Overview of Russian small cats. In: Journal of Mammalogy, Vol. 5. Berlin 1930.
Peters, H. B.: About the breeding of hybrids of wild and domestic cats. In: Veterinary Review, Volume 38, No. 51, Berlin-Wittenberge 1932.
Pocock, R. I.: On the English Domestic Cats. In: Proc. Zool. Soc. London 1907, Hälfte 1.
Exhibition of and Remarks upon two Photograph» of a Hybrid between a male European Wild Cat (Felis silvestris) and a Female African Wild Cat (Felis ocreata ugandae). In: Proc. Zool. Soc. London 1907, Part 1.
Pocock, Notes upon some African Species of the Genus Felis, based upon specimens recently exhibited in the Society Gardens. In: Proc. Zool. Soc. London 1907, Part 2.
The Races of the European Wild Cat (Felis silvestris). In: Journ. Linnean Soc. London (Zoology), lid. 39, 1934.
Rode, P.: The Use of Some Characteristics of Hair in Mammal Systematics. In: Bull. Museum of History. Nat., 2. Ser., Bd. 3. Paris 1931.
Rode, P. et Didier, R.: On a wild cat from France. In: Bull. Museum of Hist. Nat., 2. Ser., Bd. 7. Paris 1935.
Schmidt, F.: Contributions to the knowledge of the wild cat and its crossing with the domestic cat. In: Der Deutsche Jäger, 63rd year Munich 1942.
Schwangart, F.: Tribal history, race studies and breeding system of domestic cats. Published by A. Heber and Co., Leipzig 1929.
Schwangart, F.: Breeding classification and evaluation Regulation (standard) of the "Association for Cat Breeding and Protection of Cats E.V.", HQ in Dresden. Relevant as a guideline statute for the exhibitions organized by the Federation. 1st and 2nd ed., Undated (published by the Association HQ Dresden-Freital 1929).
Schwangart, F.: On Racial Constitution and breeding of the house cat. In: Journal for Mammals, Volume 7. Berlin 1932. Also published separately by the "Deutsche Ges. for Mammal science."
Schwangart, F.: The manul, Otocolobus manul (Pallas), in the system of the felids. In: About true cats and crawling cats (Felidae - Viverridae), Carnivore Studies, Vol. II, Central Journal "Small Animals and Fur Animals", 12th year, issue 8. Published by Dr. P. Schöps, Leipzig 1936.
Schwangart, F.: About our wild cat (Felis silvestris Schreber). In: The German Hunter, 59th year Munich 1938.
Schwangart, F.: South American bush, mountain and steppe cats. In: Dep. Bayer. Ak. Science Mathem.-Naturw. Abt. N. F., No. 49. Munich 1941.
Schwangart, F.: To F. Schmidt: "Contributions to the knowledge of the wild cat and its crossbreeding with the house cat". In: The German Hunter, 64th year Munich 1942.
Schwarz, E.: The wild cat of the Balearic Islands. In: Zool. Number 91 vol Leipzig 1930.
Stromer von Reichenbach, E.: The vertebrae of land predators. In: Zoologica, Bd. 36. Stuttgart 1902.
Vosseler, J.: Angora cats and angora goats in their homeland. In: The fur animal breeding, Vol. 3. Leipzig 1927.
Zorzi, F.: The boat stilt house of Solferino. In: Bull. Of Italian Palentology. N. S. Bd. 4. Tivoli 1940.
Zukowsky, L.: About two new forms of Felis caudata Gray. In: Arch. F. Naturg., Vol. 80 A, Issue 9. Berlin 1914.
Zukowsky, L.: Three new breeds of small cat from West Asia. In: Arch. F. Naturg., Vol. 80 A, Issue 10. Berlin 1914.
Zukowsky, L.: Description of the skull of Felis caudata macrothrix Zukowsky. In: Arch. F. Naturg .. Vol. 87 A, Issue 10. Berlin 1921.
LIST OF ILLUSTRATIONS
TABLE 1
Fig. 1: Wildcat from the Harz Mountains (wild caught). According to F. Schmidt, research centre "Deutsches Wild", see here under "Literature".
Fig. 2: Red shorthaired tabby, domestic cat. Photo F. Schwangart.
Fig. 3: Felis sarda Lataste male, Tortoli, Sardinia, Zool. State Collection Munich, 1935, No. 67. Complete sole-stripe on hind feet. Photo F. Schwangart.
Fig. 4: Felis silvestris Schreb. female, Prince Albrecht, label Surcin, Serbia. Zool. State Collection Munich, 1940, No. 126. Left hind foot, sole-stripe. Photo F. Schwangart.
Fig. 5: Felis silvestris Schreb. male, Prince Albrecht label Surcin, Serbia. Zool. State Collection Munich, 1940, No. 28. Left hind foot, sole-spot. Photo F. Schwangart.
TABLE II
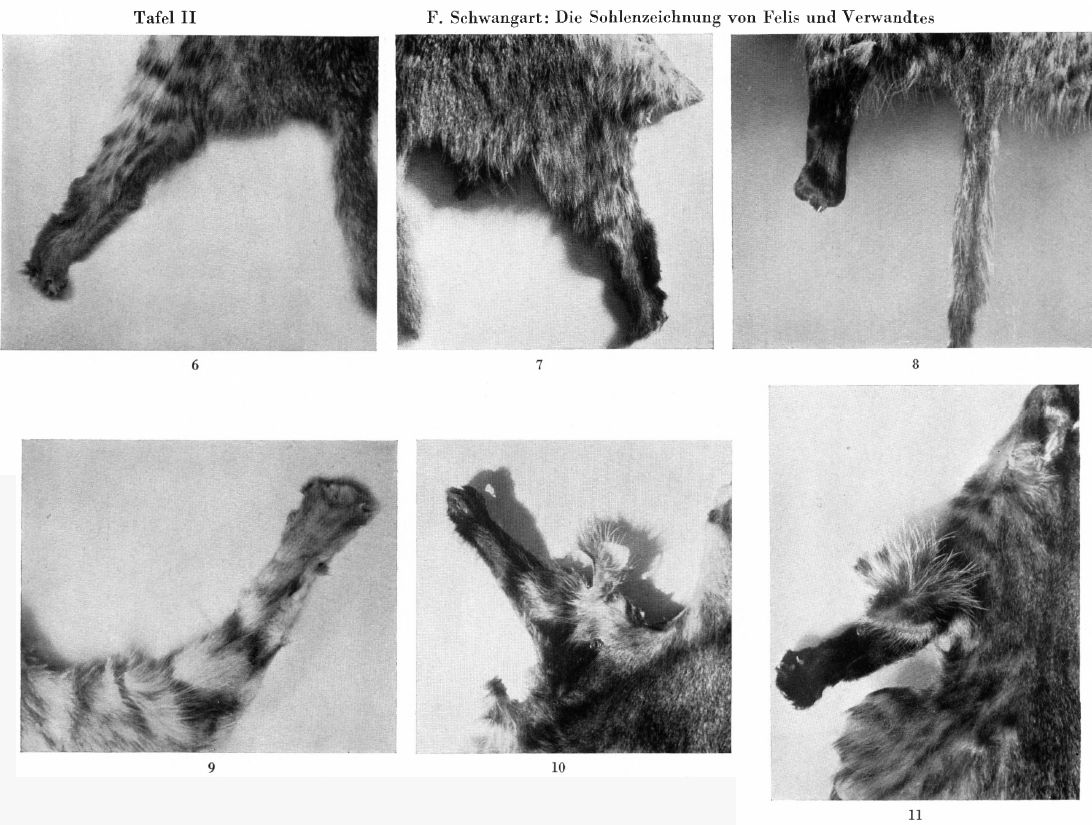
Fig. 6: Lynchailurus (Lynch.) Pajeros braccatus (Cope) emend. F. Schwangart. Dr. Schreiber and N. Jenny labelled Paraguay. Museum Hamburg, No. 42804. Colour on hind foot, phase A without blackening. Photo F. Schwangart.
Fig. 7: Lynchailurus (Lynch.) Pajeros braccatus (Cope) emend. F. Schwangart. Male. H. Krieg, collected 1931/32 in Assuncion (Paraguay). Zool. State Collection Munich. Colour on the hind foot, phase B, some blackening. Photo F. Schwangart.
Fig. 8: Lynchailurus (Lynch.) Pajeros braccatus (Cope) emend. F. Schwangart. H. Krieg, Exp. 1931/32 No. 122, St. Luis, Apabergland, Northeast Paraguay. Zool. State Collection Munich. Colour on the hind foot, phase C, full blackening. Photo F. Schwangart.
Fig. 9: Forefoot of the Lynchailurus (Lynch.) Pajeros braccatus of Fig. 6.
Fig. 10: Forefoot of the Lynchailurus (Lynch.) Pajeros braccatus of Fig. 7.
Fig. 11: Forefoot of the Lynchailurus (Lynch.) Pajeros braccatus of Fig. 8.