South American Bush, Mountain and Steppe Cats
By Friedrich Schwangart
(Treatises of the Bavarian Science Academy Mathematics and Science Department. Issue 49 1941)
With 23 illustrations in the text and 7 plates
Submitted by K. v. Frisch on 5 October 1940
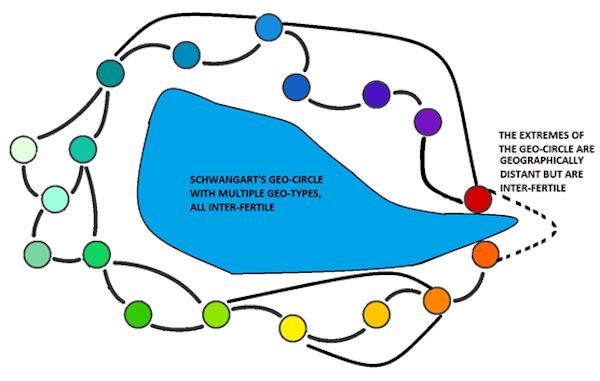
TRANSLATORĺS NOTE: A Geo-circle, or Geo-group, is a group of closely related sub-species with localised differences (unlike a ring-species, the extreme ends are inter-fertile). A Geo-type is a regional type. Modern equivalents might be landraces. Type-circles and Form-circles are used in the same way.
For a long time those dealing with the Felids have complained about the difficulties of grouping them naturally and of appropriate distinction between those in the lower categories. I mention the little known but fundamentally important criticism of H. Ihering (1910), who in turn invoked A. Brehm's judgment (from 1876), according to which the taxonomics of cats stand on shaky ground, and especially emphasizes this, for the South Americans felids. The uncertainty has not been remedied by Ihering himself, despite his basically correct requirements, due to lack of sufficient material, and not even by more recent people, including the revision made by of J. A. Allen (1919, II), which, like other authors, used only a few groups of features (the rough surface texture of the skull, some of its dimensions, peculiarities of spotted markings, whether the neck hair is smooth or swirled), where he followed the findings of O. Thomas (1903). One of the youngest felid taxonomists, Th. Haltenordt (1937), generally agrees with the complaints of difficulties in this animal family and presents a list of such statements by his predecessors. In his investigation, he confined himself to a small number of big cat species and remarked that a thorough examination of all felids meant a lifetime's work.
The difficulty in grouping these cats is an artificially created one, which Ihering likewise complained was due to the hasty increase of supposed special forms (species, subspecies, races) based on misunderstanding the geographical principles, as if the discoveries in more remote areas justified the creation of a new species (etc.), without proper knowledge of individual variability, and moreover the categories were based on minimal material, with the result that the lowest taxonomic categories multipled.
I emphasize the ecologically circumscribed groups in this work, because I have material from new forms, which I would first like to discuss, whereas the numerically larger material of the ring-spotted genera Leopardus, Margay, Oncilla is probably cause for revisions and, together with those who are thinking about relationships and about the emergence of marking patterns, for which significant contributions and changes have been made in mammals, have offered little new to taxonomy. I intend to elaborate on the problems mentioned here with reference to those genera. Likewise, in this article, the exuberant, extensively colored genus Herpailurus ("weasel cats") is omitted.
However, I would like to provide more general information, especially about the characteristics currently used and perhaps to be exploited in the future, whereby asides on the individually untreated genera are unavoidable. In addition to the markings and features of the hair, the skull has been used mainly for species and genus differentiation according to overall shape, bone thickness and some individual features. The difference in thickness of the skull walls, which, according to the treatise by O. Thomas (1. c.), especially emphasizing Allen (1919, II), coincides to a degree with the size of the skull or body, but in some cases where these are not closely related it is probably based on living conditions. The same is true of the degree of formation of the cranial bones, especially the crista sagittalis, whose stronger development may be a feature of a firmer skull, but which is also linked to greater bite power and, in different species, is also linked to age and gender. An example of the second category mentioned is the contrast between the approximately equal genera Oncilla pardinoides and Oncifelis geoffroyi ssp salinarum. According to previously known finds, the first is a forest cat, and the second is a bush cat. The heavier skull formation in this (which is otherwise a stunted form), which approaches that of its larger relatives in the genus, probably depends on the acquisition of food. Ihering (1. c.) has already pointed out examples of the connection with "food and other biological momentum" in other mammals, and to the development of large masticatory muscles. The differences among the great ocelots (Leopardus) are particularly significant according to age, and in the "weasel cats" (Herpailurus) according to sex. Here too the shape of the zygomatic arch, whether it runs flat or protrudes, and especially whether its anterior angle is rounded or forms a somewhat projecting corner, is also a frequently used criterion. The protusion and angle are certainly also functional. Variations between species herein are significant.
As scrupulously as the authors have measured the distances between corresponding skull features, they seldom paid attention to the finer construction of the skull. The usual measurements have yielded, within limits, useful yardsticks for recognising some genera, which can also be accepted when analysing the large samples I have of some species, despite the occurrence of overlaps. However, such investigations also reveal significant differences between otherwise similar genera, as well as similarities to the point of confusion between others, such as between the ring-spotted forest cat Oncilla and the aberrant-looking weasel cat Herpailurus or the round-eared, spotted oncifelis and the pointed-eared, felis-like pampas cat Lynchailurus.
Fine distinctions in skull construction have prevailed in the classification of the Felidae: then the occurrence or absence of the external pterygoid process and the pterygoid fossa, which, to my knowledge, results from A. Birula (1916); secondly, by the boundary between the Jugal, Maxillary, Frontal and Lacrymal bones, the criterion determined by R. I. Pocock (1917) and elaborated by Loennberg (1926), which has been tested on a larger number of genera and has also been used phylogenetically; thirdly, the course of the posterior border of the bony palate (whether indented, whether nearly straight, or whether projecting) and finally the presence, reduction or absence of premolar 2 in the upper jaw, which seemingly degenerates as a continuation of the reduction of the felidĺs dentition. With regard to the former feature, the South American small cats, in all of whom the processus and fossa are presen; as well as largely in respect to the third: an indentation is present (except for rare individual exceptions). These correspondences hold true, although in the grouping of other characteristics at least three contrasting groups result, the round eared with ringed or spotted markings, Lynchailurus together with Montifelis, and Herpailurus.
Lynchailurus stands out from the other South American cats and approaches the Felis group through its steeply shaped ear and overall shape, and is different from them in the shape of the palate margin. As the last editor, Loennberg, points out, the small South American cats, from Ocelot downwards, are inadequately studied with regard to the second named feature, and I too have not yet been able to catch up. Remarkably, he was eventually allowed to examine some skulls of the "smaller and larger ocelots," i.e. Leopardus and one or more of the small round-eared cats such as the Margay, Oncilla, or Oncifelis, which enabled him to conclude that their skulls bore a resemblance of the puma, and pampas cat (Lynchailurus pajeros) rewsemble those of the Old World Felis and Prionailurus, so that these South Americans correspond in the construction of the zygomatic arch and possession of a fossa or of a pterygoidi externi process, but not in the form of the palate. With regard to Premolar 2, Ihering already found its individual absence or early loss in species of various genera - he refers to pajeros and colocolo as the ring-spotted "tigrina" which is his collective name for the Margay and Oncilla. Allen (1919, II) always misses this in the smallest representative (Noctifelis), which I can confirm on the basis of having investigated only a few skulls of this genus.
Among the Old World figures, this Premolar, with some individual exceptions, is missing in the lynxes Lynx, Caracal, and in the manul, which I include with the lynxes (Schwangart, 1936), also in most cheetahs, and in individuals in different genera of medium and smaller cats. It is present in six skulls of the very small island forms of prionailurus in our collection. Its absence, presence or stunting seems to be related to the length of the jaw, and possibly also to the proportions, but in any case is not related to its natural relationship with other species. I (1.c.) examined several genera of the felids for the degree of development of a rough tuberculum on the nasal process of the maxilla, and commented on its supposed functional significance. As in most genera, it is also subject to large fluctuations in the majority of South American cats. It is noteworthy that it is consistently strongly developed in the weasel cats (Herpailurus), and in the two manul skulls, which I examined. As a sign of systematic interdependence among genera, these states are not considered thereafter.
In view of the great individual variability of felid skulls in the vast majority of pieces, to which Pocock (1916) made made a classical contribution in his studies on Scottish wildcat skulls, one can credit the discoverers of constant features, that is Birula, Pocock and Loennberg, with great merit. I have been able to fully confirm the Pocockĺs results on the Scottish wildcat in terms of modifiability by examining the distant South American genus Leopardus (ocelot). There is very rich material from the journeys of H. Krieg.
HaltenordtĹs meticulous investigations (1.c.) on large cats represent a comprehensive advance in the same area and should be followed in small cats. The uncommon degree of individual variability of parts of the skull contributes greatly to the difficulty of systematic classification of Felids, along with the oft lamented relative uniformity between species. By undertaking a classification based on skull construction HaltenordtĹs endeavours (p. 238) contrast with those of Ihering (i. C.) and give a more versatile system, a direction which I also recommend. Therefore, detailed skull examinations are not included in the scheme of this work, and are omitted in favor of other images that will follow in other contexts. As to his classification, however, Haltenordt said that he was concerned with first applying his examination method to the skulls (p.101), and that he did not regard his subsequent classification as definitive. Meanwhile, there is a critic of his methodology. S. R. Zarapkin (1939) denies the possibility of coming up with natural systematic categories, because "we can not solve this problem definitively because we lack perfect material and our knowledge about felids is still very poor."
To my knowledge work on other skeletal parts of South American small cats are still pending. Although I have found no usable features even when investigating a smaller number of skeletons of several genera, I consider it probable that we ca at least determine functionally-induced differences between genera which are already differentiated by external appearance. I would like to recommend two separate pieces of the skeleton for investigations on distinguishing characteristics: Especially the penis bone or its rudiment. And, in this connection, perhaps even more importantly the penile spines, which differs individually in the domestic cat for example, and certainly also in the felids. It is an old misery of the mammalian systematist that it is based almost exclusively on skulls, skeletons and skins. As well as organisers of future research trips, the zoological gardens could earn merit here. Secondly, the rudementary clavicle could be considered. Differences in this will probably be more functional (running cast vs climbing cats), distinguishing features only extending down to the genus. Even this piece rarely comes under our hands. From the distinction established by Pocock, basic categories based on the construction of the hyoid bone will continue to play a small part here.
There has been value in distinguishing genera and higher groups based on the coat and markings. Colouration is less important here, especially since R. I. Pocock (1907) proved it had only minor importance, something shown particularly by two groups - the Old World "golden cats" (Profelis) and the New Worldly genus of "Weasel-cats" (Herpailurus) - both of which were based on colour contrasts. The old species names "Eyra" and "Yaguarondi" have proved obsolete, and have no more meaning than the colour variants of the domestic cat. Just like Thomas and Allen, Ihering attached importance to the state of the neck hair as a generic limiting feature: whether swirled ("back curved") as in Leopardus and Margay, or smooth as in Oncilla, Oncifelis, Noctifelis. The useful feature in these confines is not used to assess the natural relationship of genera as, for example, everything else tells us that the Margay and Oncilla are closer to each other than Oncilla is to Oncifelis. Ihering wanted to use this trait for classifying the relationships of Jaguar, Puma, Leopardus and also "Felis wiedi" is which is included within the Margay. This swirled neck fur is related to the texture of the hair, also it distinguishes types above a certain body sizeM. P. Rode (1931, 1932) uses the mode of pigment grouping to distinguish between small cat species. Among the South American small cats, Herpailurus and Lynchailurus (pajeros) have proven the validity of this feature to him. According to K. Toldt (1935), the example of lynx, wild cat and "tiger cat" (Margay tigrina Erxl.), hints that differences in the form of the fur, between the wildcat (F. silvestris) and gloved cat (F. ocreata maniculata) are as good as those differences between big cats. Pursuing these investigations from the point of view of the systematic classification may be worthwhile.
The development of colours, patterns and markings has been studied in many animal groups, partly on the basis of regularities from a development-dynamics point of view (eg H Krieg (1921 and 1922, 1 and 2), Henke (1933), H. Krieg and Foerster (1937)) and partly under phylogenetics (as J F van Bemmelen, 1931), who, like H. Krieg (1922, 1, p. 90) remarked, do not fundamentally rule each other out. For the time being there is some empirical data with obvious conclusions without detailed discussion or debate.
The first question which my material has allowed me to comment on is that of the genetic relationships between solid markings and speckling to form bright rings ("rosettes"). Firstly, it was found among members of the genus Margay (Gray), that it showed a pronounced formation of rosettes in the patches on adults, juvenile specimens occur with solid markings (Fig. 1, Plate I), and there is also a difference according to sex, even in the case of the same locality, in that the females showed a stronger tendency to rosette formation (Figs. 2, a and b, Plate I). Drittens trat bei der sonst rein solidfleckigen Oncifelis geoffroyi Severtzow an einem besonders grossen Exemplar eine deulliche Hofbildung in der Schultergegend auf (Abb. 3 Taf. 2), wobei die Umrandung wie sekundńr aus Einzeltupfeln zusammengesetzt erscheint, und gleichsinnig pflegen die Hoefe bei Oncilla pardinoides (Gray), welche solche normal besitzt, an schwachen Stucken kleiner und weniger verbreitet zu sein. Bei den Lynchailurus, Severtzow (Subg. Lynchailurus) erscheinen, individuell gestuft, stellenweise Ringflecke, auch unter Kettenbildung. Die westliche Art garleppi zeigt in dieser Weise beides, die äPampaskatzen" der Ostgruppe pajeros nur die Neigung zu Ketten (vgl. die Fellbilder).
Thirdly, in the otherwise purely solitary Oncifelis geoffroyi (Severtzow), small formations of rosettes appeared in the shoulder region of a particularly large specimen (Fig. 3 pl. 2), the border appearing to be composed of individual spots, and in the same way as they develop in Oncilla pardinoides (Fig. Gray), where they are normal and less common on smaller specimens. In the Lynchailurus Severtzow (Subg. Lynchailurus) single ring markings appear in places or even form chains. The western garleppi subspecies shows both types of marking, the eastern pajeros "Pampas Cat" group shows only the tendency to form chained markings (see the fur pictures).
Thuse we may suppose that the formation of the rosette first arose from larger solid markings, it differs gradually according to sex, it "degenerates" to small solid markings (splotches, spots) and at the same time the base colour faded, this state of affairs could change in connection with the good growth rate of the individual.
If you consider the smaller forms on both sides for comparison, namely the ring-spotted genera Margay and Oncilla, the spotty Oncifelis, all "round-eared", the stippling can be associated with hunting in sunny or mostly unshaded terrain, while the ring formation is predominantly brought to completion in the deep shadow of moist, coherent forests (rainforest). H. KriegsĹ Chaco expedition clearly shows this limitation with regard to the genus Oncilla, only a single specimen of which was captured in the area of the expedition, in contrast to the Oncifelis (the salinarum subspecies) there, which he rightly called the "bush cat". The literature seems to me to confirm this mode of distribution for the two groups. The large "night eyes" of the Margay (see Fig. 1, Plate I) match the time it spends in the dark forest. Differences in the formation of the dentition, cranial ridges and other attachment points for the chewing muscles can also be reconciled with the environmental needs: The visibly stronger equipment of Oncifelis, including the thin salinarum subspecies, suits the quality of prey animals in open terrain, the thinner and more pointed snouts, especially in Oncilla, correspond to the jungle food, which probably consists mostly of tender birds. However, H. Krieg (1940) also saw the spotted Oncifelis geoffroyi in the forests of the Vorandensee Nahuel Huapi, at an altitude of 760 m, between mountains from around 1500 to 2400 m. This will be a counterpart to incidents such as those he describe (1927) for the eastern border of the Chaco area against the central and southern Brazilian complex, where flora and fauna typical of this area within the "gallery forests" spill over into the Chaco, and is described in individual cases.
The small Noctifelis guigna (Molina) seems to me to be a special case. According to the observations, which are still to be reproduced, a day animal, this small spotted cat is not completely confined to open terrain, but probably also inhabits more extensive forests. In this catĹs case, this is especially expressed by the fact that its spots are extremely dense on a very rich, almost brown background colour so that they seem almost to be absorbed. This species has its main range on the western cordillera and west of it, but also radiates east of it.
The "large ocelots", genus Leopardus, which show the transition from "small cats" to äbig cats" in terms of form, are, although ring-spotted, also widespread in scrubland, as H. KRIEGĹs yield demonstrates. Its nocturnal hunting habit ("night eyes") and body size could have contributed to the preservation of the rosetting.
Regarding those smaller forms, it should be noted that the associated rosetted ones (Margay, Oncilla) seem to be limited to the tropical area, whereas the Oncifelis occupy the entire area from the northern Chaco to the Tierra del Fuego.
To check my assumptions regarding rosettes and solid markings, additional investigations are necessary, both ecological studies and studies on a very large number of pelts, such as available in the Leipzig repository. In addition to the Noctifelis, the eastern and western South American Lynchailurus and the large and row-spotted Montifelis (nov. g.), a distinctive mountain cat, should be studied regarding their home ranges and hunting methods. In the Lynchailurus, Severtzow (Subg. Lynchailurus), as already mentioned, found rosettes appeared individually in places, and so do with chain formations. The western garleppi species shows both formations in this way, while the "Pampas cats" of the eastern group only show the tendency to chains. Compare the fur pictures.
A further question concerns the relationship between the formation of those chain spots on the basis of the rosettes, as occurs in distant forms such as Leopardus and Margay on the one hand and Lynchailurus on the other. In all cases it proves to be individually modifiable. In Leopardus, where we have samples from the same areas with a simple sequence and samples up to the fully developed "catenata" type and nowhere is there a specimen specialized in one or other of those directions, the chain formation unusable for systematic classification. In contrast, it is very suitable for studying developmental dynamics. They can be compared to the much more extensive domesticated transformation of the ancestral tiger pattern into the completely different "marble pattern" (marbleds, marbres, marmorizzati of the breeders [classic tabby]) in the domestic cat. In my monograph on Racial Constitution in the Domestic Cat (Schwangart, 1932) I dealt with this strange case and also mentioned the rare transitions between the two pattern types.
Are the pattern types of small South American cats described here an advantage for their wearer? One can imagine this from their reduction in size to speckles as well as the fading out of the background colour in species inhabiting open terrain. A causal relationship between advantage and effect is very doubtful. On the whole, van Bemmelen (1. c.) correctly states: "Looking for a physiological purpose for these secondary changes seems completely wrong to me."
Spots or stripes, which appear by coalescence of the row spots, then form chains, but these also appear anyway in the Felis genus, fore example in the tiger, so which of these two patterns is the original? According to van Bemmelen, especially on the basis of his investigations in felids, both longitudinal and transverse [horizontal] "stripes are created by lining up spots." He traces the row spotting back to the overlapping scale covering of the hypothetical ancestors. The picture with the ocelots and their smaller close relatives seems to confirm the development of the spot. It already appears uncertain with the lynchailurus. My Lynchailurus (Pseudolynx) kriegi n. Subg. n. sp. described here (Fig. 22 a-c) Plate VII, with its alternation and transition between row spotting and stripe formation, parallels the stirped domestic cat. In the case of the domestic cat, the domestically acquired stripe type, which never forms spotted rows, can hardly be traced back to that pattern. Likewise, the conditions in the small South American cats do not allow any transfer to, for example, the striping in Panthera tigris. Embryological studies such as those started by Toldt (1912) on domestic cats, where he first demonstrated the creation of the striped pattern in stained postembryo animals, could contribute to the whole question.
A systematically usable individual feature that belongs in the field of markings is, as has long been known, the white spot on the back of the ear of ocelots and their close relatives. The "weasel cats" (Herpailurus) and the lynchailurus, the pajeros group like my subg. n., lack this spot as do the related "Colocolo" (Montifelis gen.nov.). Its occurrence in Lynchailurus garleppi, albeit in a slightly different form, is one of those characteristics that, like the shape of the palate rim, seem to bridge this genus to the ocelot-like. We are still unable to estimate the value of these characteristics for phylogenetic classification.
Peculiarities of overall shape set the genus Herpailurus (weasel cats) apart from the other South American cats, especially since the lack of markings is innate. In contrast to the certain colour variants of this genus, the similarly colored puma, which was previously classed with them due to its youthful spots, more closely approximates the spotted small cats, which, according to Loennberg (1. c.), also seems to apply to it because of the type of boundaries between the pieces of bone on the front part of its zygomatic arch. To link the Puma to small cats is even more tempting because the structure of the hyoid skeleton follows that of the felines rather than the pantherines, Pocock (1917). Haltenordts also seems to agree with this in his skull examinations on several "big cats", when he says there (p. 201): "Since he" (the puma) "does not have any dentition specialization whatsoever, one could call him the house cat type among the big cats precisely because of his rounded and broadened skull design, which of course does not mean that he is somehow particularly close to Felis catus, but that characteristics of his skull are like an enlarged version of her." Perhaps a conclusion about a closer relationship or affiliation to the felines (mostly small cats) and its contrast to the pantherines (Pocock) is allowed, if one allows the inclusion of its hyoid bone apparatus shape, which is obviously an essential systematic feature, as well as features of its behaviour, including its peculiar voice, circumstances which cannot be discussed here. Haltenordt tries to reduce the systematic value of the hyoid apparatus (p. 234/35) but, to me, his reasons do not seem sound. Regarding the case of Herpailurus, it should be remembered that these cats, which seem so isolated, like Lynchailurus, again approach the other small South American cats in the shape of their palate, in contrast to the old-world cats.
The zookeeper, breeding biologist and sensitive observer of animal behavior, O. Antonius, has repeatedly pointed out the importance of the external appearance of the living animal, both at rest and in characteristic movement, for a natural systematc classification. Often these criteria provide reliable ways of differentiating where skull and pelt zoology fail. I am convinced that many features of behavioural features, many psychological expressions, can give systematic-phylogenetic hints, as much as one has become accustomed to only and always basing systematics on ethological-ecological conclusions. One can make the same distinction in such phenomena - between old traits and more recent acquired ones - as in the case of somatic formations. (As I have demonstrated in hte case of tail movements in Lynx and Otocolobus manul, 1936.) The application of such standards to the small cats would be greatly encouraged, as soon as our zoos have resolved the issue of their natural accommodation and our explorers must consider behavioural observation equally valuable while collecting data, rather than maintain a purely utilitarian standpoint. In zoos one could also assist with the difficult cat systematics through serology and through attempted hybridization - whether produced at all, and whether fertile.
The superiority of external appearance as a systematic yardstick over features such as the skull is often unmistakable in small cats. If, according to the skull, the "Pampas cats" (Lynchailurus) are difficult to separate from the spotted Oncifelis, the "weasel cats" (Herpailurus) from the rosetted oncillas, these genera contrast significantly in both cases by their head and face shape. Compare the general constitution pictures of "Tiger cats" (Fig. 1, plate I and Fig. 11, plate IV) with that of "Pampas cats" (Fig. 5, plate II and Fig. 15, plate IV). Thus the complete picture of a "Felis colocolo" Molina given by Philippi (1870) proves that, despite its shortcomings, this cannot be an Oncifelis, as Allen (1919) suspected. Such differences often appear to be phylogenetic rather than ethological.
Ihering, who also paid attention to this kind of distinction, found that among the ocelot affinities the smaller species had a shorter face than the larger ones; thus "tigrina" (in his mixed term for Margay + Oncilla) had a shorter one than Leopardus. After a more precise division into genera has taken place, this can no longer be maintained: the narrow heads of the small oncillas appear more pointed than those of the larger Margay tigrina, the diminutive form Oncifelis geoffroyi salinarum has the same relatively short-looking snout as the typical large Oncifelis geoffroyi, etc. Here, skull measurements have to play a role in order to help determine which distances cause the impression. A large part of the external shaping depends on the muscle. Ecological factors play a part in this, depending on the type of prey and prey capture in a certain area, but this is precisely where our knowledge lets us down.
Conclusions can be drawn from the form of the eye. That of the Margay fig. 1 plate I impresses us as a typical "night eye", that of Lynchailurus fig. 5 plate II as adapted to daytime, that of Oncifelis fig. 4 plate II is probably intermediate. As far as we know, the Margay is an animal of the closed forest, Lynchailurus a "pampas cat"; the Oncifelis evidently prefer bush terrain like that in the Chaco. From his travels in this area, H. Krieg brought us a whole collection of the Oncifelis geoffroyi salinarum form, but, as mentioned, only a single rosetted specimen of the genus Oncilla, which is close to Margay, from the area of Villa Hayes, about 80-90 km north from Assuncion (Paraguay). For this isolated occurrence I received an interpretation from an expedition member, the painter Kiefer-Munich, which I would like to cite as plausible in this context. The Rio Paraguay separates the Chaco side with its typical vegetation from a fertile zone of red soil with lush vegetation and medium-high tropical forest. From there, large animals from the wooded side can also come to the so-called floating islands of the Chaco side, to Leguane. Caimans also float across in this way. The reporter now suspects that isolated forest cats, such as the oncilla here, reach the Chaco area in this way, which they otherwise avoid.
A feature of uncertain importance for the systematics is the shape into which the pupil contracts. It was thought to be purely dependent on the hunting season (day, twilight, night) or the way of hunting (a more horizontal or more vertical view), as Ihering says, "more on its way of life than its physical organization", but data on the behaviour of different cat species does not support this interpretation. The extent to which the South American small cats have "slit pupils" has not yet been decided, at least not for Lynchailurus. In a special study, K. Schneider (1930) has shown that among the Carnivores there are intermediate and otherwise deviating pupillary shapes, also changes according to age (e.g. in the tiger). Among the "small cats", to which slit pupils were ascribed, the Manul has an almost rounded constriction.
R. I. Pocock (1917) published a study on the shape of certain externally visible parts of the body in the Felidae and particularly pointed out the systematic value of the ear shape. In the case of the small South American cats, the following can be considered: The ocelot relatives (Leopardus, Margay, Oncilla, Oncifelis, Noctifelis) with relatively large round ears, like those of the old-world spotted cats and relatives; - the weasel cats (Herpailurus) with the smallest auricle of all felids, which is also rounded, but in its somewhat tall form cannot be classed with the latter (Fig. 6, plate II); - the steppe cats (Lynchailurus) and the mountain cat "Colocolo" (Montifelis mihi) have an upright ear, which is similar to that of the genus Felis. Although these opposites can clash with an approximation of previously mentioned, systematically used features - the typical "South American" shape of the palate rim, e.g. also in Herpailurus - one can hardly avoid the impression that they are of particular tribal historical value. If it is often difficult to determine the shape of the ears on skins and pelts, this applies even more to the shape of the rhinarium [nose leather] and the form of the claws, which Pocock, as director of the London Zoo, was able to get to some extent. He found that the rhinaria of different genera of the ocelot group to be the same (Leopardus, Margay, Oncifelis) and, as expected, different from that of Herpailurus, whereas surprisingly Oncifelis g. geoffroyi like g. salinarum merged with Herpailurus in contrast to Leopardus and Margay. Ihering found general differences in the number of teats, but claims to have "only used them with caution" himself. He also suggested paying attention to the characteristics of the penis, which as far as I know no-one has done in terms of systematics and which I have already spoken about here.
From what I have said so far, it is clear that it is desirable to establish the systematics of the South American small cats and the Felidae in general on a more multi-faceted basis than has previously been the case. I have also already commented on the preconditions for future procurement of suitable research material and their implementation.
Require attempts to derive the genealogical data of these small cats requires knowledge of prehistoric conditions. N. Kretzoi (1929) in his special paper on the fossil Ailuroidea states that they were absent in South America until the Pleistocene. At the beginning of this era some migrated there via Central America. There were the smilodonts (machairodonts), which have no connection to the modern felids, and some felid genera that came from Asia to North America, roughly on the line of the Bering Strait. There are no extinct representatives of the current genera of South American small cats (which does not exclude the possibility of finding some), but there are representatives of two extinct species of the current genera of large cats, jaguar and puma. I have mentioned the resemblance of the puma, based on the structure of its hyoid bone, its facial structure, and features of its behaviour, to the small cats, and R. I. Pocock's decision to add it to this main group; whereas, based on his skull studies albeit they have not yet extended to small cats, Haltenordt places it in the panthera genus.
Jaguars are probably ruled out as ancestors. The ocelot relatives can most likely be traced back to puma-like ancestors, whose representatives, as far as Loennberg (1926/27) examined them, also resemble the puma in terms of the positional relationships of the pieces of bone in the front part of the zygomatic arch. The pointed-eared, felis-like Lynchailurus, together with the Colocolo, are further away from it in their overall conformation - even after details such as the positional relationships of those components of the zygomatic arch -; likewise the Herpailurus' (weasel cats). These appear "primitive" or Viverrid-like. I think one should not completely ignore this striking similarity, even if Kretzoi (1. c.) believes in convergence based on paleontology. But Haltenordt even applies the comparison to the lion by calling him a "Viverrid type among the big cats" because he "exhibits so little of what is commonly understood to be cat-like".
A derivation of Lynchailurus directly from the Felis type or Asian closest relatives, comes up against the contrasting form of the systematically utilized posterior edge of the palate, but there is also accordance on this point with the East Asian genus Prionailurus, whose southernmost representative are cats of the Sunda Islands (Bengalensis group ), the northernmost of which is the "Amur cat" (Pr. euptilurus) (these two groups should form at least subgenera). The Amur cat is characterized by a raised, dome-like ear and also in its overall structure - shape, face shape, details ľ it resembles Lynchailurus, as well as Felis, in its spotted type it somewhat resembles the contrasting spots of Lynchailurus west of the Andes (garleppi). For the ocelot group (Leopardus, Margay, Oncilla, Oncifelis, Noctifelis), apart from the interpretation of the puma relationship, there also remains the question of Prionailurus, namely with the round-eared Bengalensis type, which even has the light ear mark. Perhaps the ocelot-like lynchailurus, on the other hand, is due to various waves of immigration in the Pleistocene, whereas the "aberrant" Herpailurus is a branch of Viverrids remains conceivable. Anatomical studies would have to contribute to the decision about the Viverrid character of the Herpailurus. Some fossil viverrids have long been known from America itself (Aelurotherium Adams 1986, see Trouessart, Catalogus Mammalium 1904).
In the descriptions that now follow of some new and revisionary remarks on described South American small cats, I mention the peculiarities of the skull only with reference to criteria that have already been given. The skull was only available in one case for the new descriptions, fortunately these animals are largely characterized by peculiarities of their coat pattern. One often misses a portrait very much; in several cases photos of living animals from the Vienna-Schoenbrunn Zoo (Prof. Dr. Antonius), the Berlin zoo (Dr. Lutz Heck), the London zoo (Mrs. Prof. Veley(London)) and one of the excellent drawings by Prof. Dr. Krieg have helped. Measurements of skins often only provide uncertain contributions to our knowledge of the proportions, due to deformation caused during preparation. The terminological starting point was the work of Allen (1919, II). From his work (1919, I) I accept the foundations of M.N. Severtzow's (1858), the admissibility of which has been repeatedly criticized, but which, despite the lack of descriptive justification promised by the author but not carried out, are identifiable and show a significant systematic view.
My material from the genera I will mention here, such as from the "rosetted" small cats and the genus Herpailurus (on which a publication is to follow after complete processing of material), came from the museums in Munich, Berlin, Hamburg, Stuttgart, Genoa (supported by informational letters from the director Dr. O. de Beaux), whose contributions I examined here on site, and from Paris, Milan and Turin, where I stayed for the investigation and received support.
I would like to thank our taxidermist E. Schuhmacher for many of the photos made here, and for Prof. Dr. Antonius-Wien-Schoenbrunn and Dr. Lutz Heck, Berlin Zoo. The Berlin museum only sent skins and hides, not skulls, as it wanted to reserve the skulls for HaltenordtĹs planned special investigations into small cat skulls.
Oncifelis Severtzow 1858
Oncifelis M. N. Severtzow, Rev. et Mag. De Zool. Ser. 2, vol. 10.
Herpailurus R. I. Pocock, Ann. Mag. Nat. Hist. Ser. 8, vol. 20 (1917). Partim: Without Herpailurus Severtzow, 1. c.
Oncifelis, J.A. Allen, Bull, of the Am. Mus. of Nat. Hist. Vol. 41 (1919). Partim: Without the species Colocolo (Molina).
The animals of this genus are "spotted cats" with fine or coarser spots, which, otherwise monochrome, in Oncifelis gracilis n.sp. can consist of two shades. Their skull structure shows traits of robustness, as demonstrated by Allen (1919), e.g. In some cases based on older descriptions, - relatively significant zygomatic width, massive walls, well-developed sagittal crests, strong teeth, this also applies to the small salinarum form, which means that its skull can also be consistently differentiated from that of the slimmer, more finely built, rosetted Oncilla type. The difference in shape becomes even clearer when comparing the heads of living individuals; see. Fig. 1 plate I of a Margay and Fig. 4 plate II, Oncifelis geoffroyi.The Oncilla's face will look even slimmer than that of Margay, according to the structure of the skull.
According to the proportions of the bulla, Oncifelis geoffroyi switches between the rosetted cats on the one hand and the genus Lynchailurus, represented by L. pajeros, on the other - R. I. Pocock (1917). In the first-mentioned group, the inner chamber takes up by far the most space, in the latter the outer chamber takes up more space. I find considerable fluctuations in all three, which makes a demarcation of Oncifelis on both sides impractical on the basis of this standard, even if the distinction between collections applies to the majority.
In material of the genus has two species, of which one, geoffroyi, has three distinguishable forms. Each of these obviously has its own geographical focus. Their systematic value is different and the connections between them remain uncertain due to the lack of larger collections, one has to limit oneself to guesswork.
1. Oncifelis geoffroyi (dĺOrbigny and Gervais)
Felis geoffroyi dĺOrb. and Gervais, F. E. Guerins Magazin de Zoologie, II. Ser., 5th year, 1843. F. geoffroyi dĺOrb. and Gervais, Voyage dans lĺAmer. Meridionale, 1847. - Felis (Oncifelis) geoffroyi M. N. Severtzow, Rev. et Mag. De Zool., II. Ser., Vol. 10 (1858). - Oncifelis geoffroyi (dĺOrb. and Gervais) J. A. Allen, Bull. Amer. Mus. Nat. Hist. Vol. 41 (1919). - Felis salinarum O. Thomas, Ann. Mag. Hist., Ser. 7, vol. 12 (1903). - Oncifelis salinarum (Thomas), Allen, 1. c.
The original description by d'Orbigny and Gervais (1843) is based on three specimens of typical spotted cats that were brought in in 1831, two from the "banks of the Rio Negro" (Patagonia), one from "Buenos Aires". The image of a skull (1847; is probably the best of this species so far, it shows particularly well the massive structure and the wide projection of the zygomatic arches. The portrait 1843, Pl. 57, shows a distinctly grey, albeit with a light yellow, background hue and a relatively wide distance between the spots, that of 1847 has a deep yellow background hue and narrower spots. Allen (1919) found a considerable range of fluctuations in the basic hue of the fur, from "almost pure grey to pale yellow", the second hue being mentioned in the illustrations of the first person describing it. As you can see, this is not true insofar as these already show two variants, one being deep yellow and the other grey with a slightly yellowish tinge. When I was at the Paris museum, I was given the task of examining a large number of pelts and skins from South American small cats within a short period of time, and unfortunately I missed those that were stuffed. According to Dr. P. Rode (Paris Museum) one of the two "Patagonian" specimens (No. 92) is considered a holotype, the other two specimens (No. 90 from Patagonia, No. 91 from Buenos Aires) are considered paratypes. The same contrast can be found here in the basic colour and spot size as in the pictures mentioned: In the two Patagonian specimens it is unanimously a bleached white with a very pale yellowish tinge, whereas the Paratype No. 91 from Buenos Aires has a deep yellow background colour with "much smaller and more clearly arranged spots, especially on the back" (P. Rode, by letter). Allen (1919), in his summary based on specimens available up to then, limits the distribution of the species to the area between the Strait of Magellan to the south, the foot of the Andes to the west, and the upper Parana to the north where, however, no border can be drawn against the small Oncifelis salinarum (Thomas), which he and this author artificially separate.
In the following distinction between the two forms a) and b) of Oncifelis geoffroyi in terms of their first descriptor, due to the difference in colour and other coat characteristics, the identity of these two had to be taken into account with the variants identifiable in dĺOrbigny and Gervais, according to the types and the images. A decision about this could only be made by asking Dr. Rode (Paris), as additional features, such as the hair length and the colour of the soles, are not recognizable in the pictures and are not mentioned in the information I have received about the types. These features are clearly evident in the three silver-grey specimens from the south in Munich. If they are just as pronounced on the Paris holotype No. 92 in contrast to the paratype No. 91, then the holotype (number 92) named Oncifelis geoffroyi geoffroyi dĺOrb. and Gervais 1843 has to be considered fundamental and the name Oncifelis geoffroyi (dĺOrb. And Gervais) forma argentea Schwangart, which I have given for the time being, should be withheld until verification is possible. Even if the same were true only of paratype No. 90, it would not be the deep yellow variant (like Paratype No. 91, 1847), but the grey variant, according to the data in the illustrations. It is possible, however, that the grey Parisian specimens compared with the Munich specimens represent transitions to the deep yellow form.
a) Oncifelis geoffroyi (dĺOrb. and Gervais), forma argentea mihi.
Felis geoffroyi dĺOrbigny u. Gervais, holotype no.92, paratype no.90, Mus. Paris. - F. E. Guerins Magazin de Zoologie, II. Ser., 5th year, 1843, fig. 57.
The following description is based on three skins from the Munich State Collection, which are to be regarded as cotypes of this form (1930 No. 134, 135, 136, see Fig. 7, plate III). They belong to the Otto Becker collection and have the location Comodoro Rivadavia, a port in Chubut. They differ from all of the skins of both the larger variant (to which O. Thomas (1903) and Allen (1919) restricted the name geoffroyi) and from the smaller variant (Thomas's salinarum) which I saw, because of their distinctly silver colour, hardly yellowish background colour, their somewhat longer and especially softer hair - including variations in the other skins - and an almost purely black "sole stripe" covering the entire sole, while this is either distinctly brown in other skins or is replaced by a series of shadow spots. The spots on the trunk are unequal in size in the three skins, their distances are wider than in other skins of the species. There are no skulls with these skins.
The dimensions are remarkable. The largest piece has a total length of 110 cm, the second largest being 102 cm, which means that both fall within the range of AllenĹs Oncifelis geoffroyi. The smallest, however, measures only 87 cm, with no signs of being a juvenile. According to its measurements it falls within the range of Thomas's Oncifelis salinarum, which was described by its author as a "small northern representative of F. geofffroyi" and was only known in different shades of yellow. We shall come back to this when discussing that form.
The Rio Negro, from whose banks the above-mentioned yellowish-grey type specimens of d'Orbigny and Gervais originate, forms a dividing line between two types of terrain, the lowland, which leads northwards to the pampas reaching as far as the plateau of Mato Grosso, and the extensive Uttak Mountains, to the south adjoining the hill country of Chubut, traversed by the river of the same name. Since the three Otto Becker skins in our collection come from there, it can be assumed that this is the typical form limited to the southern Patagonian landscape remote from the area of the yellow backgrounded larger and smaller geoffroyi and that it represents a geotype there, which may of course be temporarily connected with the more northern yellow representatives. It remains to be seen which group the Oncifelis geoffroyi mentioned by H. Krieg (1940) on the Vorandensee Nahuel Huapi, headwaters of the Rio Limay belong to.
b) Oncifelis geoffroyi (dĺOrb. and Gervais), forma flava, mihi.
Felis geoffroyi dĺOrbigny and Gervais Paratype No. 91, Mus. Paris. - Voyage dans lĺAmer. Meridionale, 1847, fig. 14.
These are the larger, yellow-backgrounded pieces that I used as templates. The illustration in dĺOrbigny and Gervais 1847 shows the closely spaced stippling perfectly. Of two Parisian skins, one with the designation "Felis geoffroyi, Argentine" corresponds to the colour and size according to the pattern which Allen (1919) regarded as typical geoffroyi and is to be regarded as a paratype of the form; the other, smaller, is infantile according to the skull associated with it and is probably to be placed here. The Berliner Pelt No. 13278, male "Paraguay, Berlin Zoo", is somewhat overstretched with its total length of 116 cm, but is inherently the largest piece that I have encountered. In the shoulder region and behind it, is the ring formation already mentioned as an exception in this genus (Fig. 3 Plate II), which here, in contrast to the rosetted genera (cf. Fig. 2 Plate I), appears to be coalesced spots; otherwise this pelt is very spotty. Base colour is tawny yellow, the back is brownish. On the anterior part of the torso there is a degree of horizontal arrangement of the spots (Fig. 8, Plate III). The Hamburg material, "F. geoffroyi d'Orb. Female, Zool. G. December 2, 1891, Buenos Aires, E. K. 22276", with skull no. 22277, shows particularly well that there is no consistent difference in the extent of the spots compared to the smaller form ("salinarum"); it fits exactly into H. Kriegs "Bush cats" (see below form c). The background is deep yellow. Whole length is about 100 cm against 94 of the longest skin of the others. From here, as well as from Otto Becker's silvery skins (Forma argentea, see above), size transitions lead to the Salinarum type. The skull differs only in size from the others, its common oncifelis features (Allen) are less pronounced (female): the anterior corners on the zygomatic arch protrude only slightly, the arch itself bulges relatively slightly, occipital crest is relatively delicate. The bullae also appear slender and thin-walled on the oncifelis scale. Also due to the development of the onciform features of the skull in comparison with the following, as required by Allen, a consistent delimitation cannot be achieved.
c) Oncifelis geoffroyi (dĺOrb. and Gervais) forma salinarum Thomas.
Felis salinarum O. Thomas, Ann. Mag. Hist. Ser. 7, vol. 12 (1903). - Oncifelis salinarum (Thomas), J. A. Allen, 1. c. 1919.
Pocock (1917, External Characters) and Allen (1919) already have reservations regarding the species-establishing value of the characteristics cited by Thomas, in opposite directions: Pocock considers salinarum a subspecies of the larger "Oncifelis geoffroyi", but Allen would like to insert it as a subgenus between the spotted genera Oncifelis and Noctifelis. However, his own pelt photos show it should be placed close to the larger representatives of the genus and differentiates it from Noctifelis, which I doubt, incidentally, should maintain genus rank compared to Oncifelis. Allen limits the distribution of salinarum to southern Bolivia and northern Argentina, reaching altitudes of 8700 to 9400 feet. The name "salinarum" alludes to the "often very strong soil salinity" in a large part of its distribution area.
The basis of the material that was available to me is the H. Kriegscher collection "Bush cats", 8 pieces, locations: Chaco (Mennonite area), Villa Montes, upper Pilcomayo, three pieces bought in Assuncion. For "Exp. 1931/32 No. 48, Chaco boreal, Mennonite area", skulls and skeletons are present, as well as two skulls without skins (1926 No. 606 and 1926 No. 653 both Villa Montes). As far as is known, the sites are in the Chaco plain and near Villa Montes on the western edge of the Cordillera. Fig. 9 Plate III shows the type of spots on one of the skins.
The basic colour of all these skins is pale yellow, that soon has a more brownish, now a somewhat grayish tinge. The markings show the pure dot type without any signs of rosette formation. In some pieces, a tendency to horizontal orientation of the spots is noticeable (see Fig. 9, Plate III), as I have also described for one of the large geoffroyi pelts. One finds a weakening of the marking impetus in this lax orientation, in harmony with the stunted spots, rather than any phylogenetic hint that would contrast this species with the rosetted genera and a pronounced tendency to change towards row-formation. The hairs are short and quite hard with minor deviations that are somewhat softer. The longest pelt is 94 cm, of which 35 cm is the tail, the lengths of the others range from 76 to 90. All pieces can be considered adult. Regarding the skull, I limit myself to pointing out that the dimensional differences in these few pieces of a collection already show the uncertainty of some of the limiting features given for the genus Lynchailurus, as the appearance of the head and face of the two genera is fundamentally different. This subheading includes the shape and size of the bulla, the ratio between its chambers, the width of the palate region, dimensions on the mesopterygoid, and the allegedly rather more delicate dentition in Lynchailurus. As the greater material confirms, these distinctions apply only to the majority on both sides.
I also had:
Two Parisian skins "Felis mitis Fr. Cuv., Chaco", a juvenile specimen "F. geoffroyi, Chaco ". - From Turin: pelt no. 1529 with skull no. 4057 "F. geoffroyi dĺOrbigny, Pampas centr. Argentina, Dono. Dott. Giov. Bertoldo" and No. 535 "F. geoffroyi dĺOrb., Cala, Argent.", with skull 2282 "Dott. Alfredo Borelli"; all typical of this form. - From Genoa: a hide and two skulls that do not belong to it, all from "Patagonia"; the coat "Oncoides geoffroyi dĺOrb., ded. Dr. de Beaux, Rio Negro ", i.e. from the area where skins 92 and 90 were found by dĺOrbigny and Gervaisĺ original description; both skulls "Oncoides geoffroyi dĺOrb. (?), Patagonia, sped. Ant. It. G. Bove 1884, C. E. 17635 and 17636", with the note "Etichetta originale sperduta Dr. de B." The first-mentioned skull appears abnormal, the second very much resembles the above-mentioned by H. Krieg (Exp. III No. 48) as a typical of a small adult Oncifelis, belonging to the salinarum type. The same applies to the Genoese fur, the length of the trunk (in the absence of the tail) being 60 cm, so that its total length fell into the "Salinarum" range, as evidence of the occurrence of this type on the border with Patagonia. A stuffed Genoese piece that was assigned to Oncifelis, according to a letter from the director Dr. de Beaux also came out as belonging here. - From Hamburg: Skull 38360 "Felis macrura Wied, Paraguay, L. des Arts March 9, 1905, an Oncifelis assigned in size to the "bush cats" H. Kriegs, more compact and massive than that of the already discussed piece of large form, No. 22276/22277, male. Another skull, no. 42669 "Felis geoffroyi male juv. Zool. G. 25. 11. 1929", has infantile features in its smallness, the pear-shaped bulging of the roof, the particular shortness of the snout is an infantile characteristics and, in view of the relationship between the size and the tooth changes in progress, it appears retarded and abnormal. The oncifelis-like zygomatic width and corner formation in front of the zygomatic arch stand out clearly, while the crests are hardly suggested. A safe allocation to one of the forms is not possible. - The Berlin material (pelts only): No. 7179 female "Tornquist Buenos Aires P. Neumann S.G.", whole length 98/99 cm, and No. 7178 male "Tornquist, at Bahia Bianca, Buenos Aires, P. Neumann S.G.", full length 90 cm, perhaps not yet fully grown, has colouring and spot characteristics close the "bush cats" from the Chaco on the one hand, while the Berlin piece 13278, on the other hand, because of its size, should be placed with Oncifelis geoffroyi forma flava. The specification Bahia Bianca defines a southern Argentine coastline of predominantly pampas character as the area of discovery. No. 21319 "Oncifelis, Casca, Pilcomayo area, Bolivia, W. Herman" is a small, very pale yellow fur that follows those of H. Kriegs in its markings, but with its reduced speckling looks particularly poor. No. 21321 "Tanja, Pilcomayo area" etc. stands in considerable contrast to the latter due to its rich yellow base colour with striking, conspicuously large, almost black spots that show a tendency to form small hooks and horseshoes from the front to the central back, but not to form rings. With the exception of the small size, total length 91 cm, it does not give an impression of a stunted form.
The overall impression created by this representation of the salinarum form can be summarized as follows: It seems that a region of its exclusive occurrence has developed, the northern Chaco, the northern Chaco, where it lives in the lowlands, but also reaches heights of up to about 3000 m (Sucre, Bolivian province of Chuquisaca, according to Allen) further down. Specimens of this form can be found further south in the same areas as the larger yellow form, and this distribution extends southward to the adjacent region where the Munich silver fur (O. Becker collection) occurs. As already said, one of these skins resembles salinarum in spotting and size. In terms of size there are transitions, but according to the structure of the skull and the type of spotting there are no fundamental differences between the larger geoffroyi and the salinarum form.
Oncifelis salinarum (O. Thomas) subsequently appears as a variant occurring in large areas among the forms belonging here, usually alongside the appearance of a "stunted form". Circumstances which would have caused real concern are conceivable for the salt steppes of the Chaco as well as for mountain areas; at least in the first-mentioned area the form occurs in isolation. [With regard to its occurrence in the Chaco, my view of Oncifelis salinarum as a "stunted form" agrees with that of the Chaco itself as a biological "impoverished area", which H. KRIEG (1938) asserted. "A lot of animal species", he states, "which occur or are common in its peripheral areas, are absent within it or decrease more and more towards its dry interior. Lots of animals . . . find it impossible to live in the low-precipitation Chaco interior. As far as they are sensitive to low temperatures or rapid temperature drops, they lack wind and cold protection. . . In the case of Oncifelis salinarum, it was necessarily resistant to these rigours, but staying in the area was at the expense of physical development. Salinarum also provides an example of the generally valid perception of Kriegs: "The special forms of adaptation that can be found in some of its landscapes, up to the salt pans, is not specific to the Chaco, but can also be found in corresponding landscapes outside the Chaco."] The poorer evidence from further south does not allow a judgment for the region. Breeding experiments in the zoo could provide information about whether the shape is hereditary in the area of its sole occurrence.
In any case, neither its distribution nor the characteristics asserted for the morphological delimitation accord it a claim to full species status. Due to its sole occurrence in a large part of its area, however, it is to be expected as a geo- or ecotype for the time being, which is to be identified by special designation. After that, the argentea and flava forms of Oncifelis geoffroyi should be added as the third Oncifelis geoffroyi (dĺOrbigny and Gervais) salinarum form (O. Thomas).
2. Oncifelis gracilis n. sp.
Furs 1934, No. 55, 56, 57, the Munich State Collection; bought from the Reiss fur company in 1934. Specification "Buenos Aires" - Hamburg Museum No. 42571 "Felis geoffroyi. Zool. Garden, July 2, 1929, female". Fur and skull. - Holotype (fur) Munich 1934, no.55 (Fig. 10, plate III), paratypes (fur) Munich 1934, no.56, 57 (fur and skull), Hamburg, as above.
The type of spotting in these pelts distinguishes them significantly from the small cats that otherwise belong to the genus Oncifelis. The spots themselves are larger, without forming rosettes and in the form of splashes, but are more often composed of two shades of colour, as in the case of rosetted genera the centre and the border may contrast. These two colours stand out not only against each another, but also against the light-to-deep yellow background colour of the skin, as is always the case with rosetted skins, but to a much lesser extent with the centre of the markings. The lighter of these two colours, which are differentiated from the background colour and from each other, is a deeper red to yellow-brown and sweeping wing-like symmetrical figures on the shoulders and sides of the neck, whereby it can merge into the darker stripes, non-symmetrically. The darker component of the spot is a deep brown. In the flank spots, this brown forms a point facing towards the tail, while the lighter component forms a broader, head-facing basal portion. In the overall impression, the pelts appear more or less foxy, as in no other South American cat known to us. Markings on the back, interrupted multiple stripes, are continuations of the dark neck and head stripes, which in the classification system resemble those of the other rosetted and spotted South Americans cats. The entire torso pattern is decidedly longitudinal. The tail - only complete in one of the Munich pelts - has irregularly arranged ringlets that disintegrate into spots. The legs from thigh to foot have black-brown spots; the irregularly delimited sole spot is only preserved in one of the skins. The back of the round ears have the familiar whitish marking. Neck hairs have no swirls. In terms of dimensions, this species is one of the smallest felids, it contests this rank with South America Noctifelis; its figure is likely to be more delicate and slimmer than that of Noctifelis. The fur is softer than that of Oncifelis geoffroyi flava and is similar to geoffroyi argentea.
Pelt measurements: Munich 1934, No. 55 (holotype): body length 64 cm, tail missing; 1934, 56 (Paratype): full length 82, tail 23 cm; 1934, 57 (Paratype): tail missing, end of trunk slightly truncated, trunk length about 60 cm; Hamburger pelt (Paratype): 56, tail 14 cm; infantile.
On the skull of the Hamburg specimen (paratype) the change of teeth is about to be completed; Pm 2 is missing and would have remained missing, as is regularly the case with Noctifelis. The somewhat distended brain capsule and the excessively short part of the face appear infantile. The strongly protruding and thickened front corners of the zygomatic arch point to the genus Oncifelis. Since the arch itself does not extend so much, these corners appear strangely pointed. Crest formation not (yet?) Developed. Bulla high and quite wide. Individual dimensions of the skull, which, of course, reveal little in view of its juvenile condition: total length about 71 mm, basal length 64, zygomatic width 48+, postorbital narrowing 27, postorbital processus distance 37, molar distance 30, hard palate length 28+, canine distance about 17.
The species, with its splotchy, sometimes two-coloured type of spotting, could lead into the rosetted genera (Oncilla), and I was initially inclined to assign it to that, for example as a subgenus. I decided to assign it to Oncifelis because of my acquaintance with the skull, which, despite being a juvenile, already shows distinctive features of the broad and relatively massive structure of this genus. Further material, especially the sight of a living animal, would be necessary to establish a subgenus. For now, in any case, I follow the view that under this generic relationship a merging of existing sub-genera would be more desirable than an increase, if one takes into account the requirements that are otherwise generally needed for sub-generic separation.
Origin of material from South America is likely to be certain. The seller received the Munich skins with a "Transport from Buenos Aires". There is no information about a more precise distribution area.
Noctifelis Severtzow
Noctifelis (subgenus) Severtzow, 1. c. 1858, for Felis guigna Molina.
Herpailurus R. I. Pocock, 1917, partim.
Noctifelis (genus, with Severtzow) J. A. Allen, 1919.
Noctifelis guigna (Molina)
Felis guigna, Molina, Saggio sulla storia nat. del. Chili, 1810.
Felis guigna, R. A. Philippi, Arch. Naturg., Year 36, vol. 1 (1870), year 39, vol. 1 (1873) (Fig. Of habitus and skull).
Noctifelis guigna (Molina), J. A. Allen, 1919.
Non Felis guigna Molina, R. Hensel, "Contributions to the knowledge of the mammals of southern Brazil", 1872, - which Allen includes here.
"The Guigna or Huigna" according to Molina, "wina" according to Allen and the Chileans. Allen's statement that this small cat forms a particularly sharply characterized species of spotted cats, despite the relationship between its pattern of spots and that of Oncifelis geoffroyi - which prompted H. Burmeister (1879) to adopt a local form - is best confirmed by the illustration and description of a single Zoo specimen by A. Antonius (1932); I have reproduced the picture here (fig. 11 plate IV). The large-earedness emphasized by all is clearly evident in the picture, as well as the Oncifelis geoffroyi-like but peculiar facial shape. In the text, Antonius describes the particularly tightly placed spotting, which has no tendency to any specific orientation, and the somewhat shaggy character of the hair, These features are uniform in the specimens I have used as models, as far as they are not obscured by the melanism common in this species. On the other hand, in Antonius's specimen alone, the otherwise deep brown tone was replaced by "stone grey" - perhaps a consequence of life in captivity? A second living specimen that Antonius knew, still young in appearance, whose photo I have seen, went under the designation of origin "Deseado Hinterland, Patagonia". If this is the case, then the previously known distribution area must be expanded significantly. I will return to this. The illustration in Philippi (1873) misses several features.
Noctifelis has a "sole stripe" on the hind feet, but it only differs slightly from the relatively dark background colour and fluctuates in intensity. A special characteristic is given in the double length of the tail. There are pronounced long and short tails; intermediate stages are exceptions. The meaning of this difference is explored below.
Even the skins reveal that this species is compact and somewhat low on the legs. O. Antonius saw his living specimen of "Noctifelis" "as much a ground animal as a day animal", which also more or less applies to other spotted cats, in contrast to the genus Margay; I recall the comparison of the eyes in Fig. 1, Plate I with those in Fig. 4, Plate II. The climbing tree was rarely used by AntoniusĹs specimen.
My material: 1). Skull. - No. 40350 of the Hamburg Museum. "Collection of Capt. Paessler, 3. 1920, Chile. "It resembles a smaller parallel to that of an Oncifelis. - Hamburg Museum No. 49974 "Felis guigna Mol., Ded. M. Gleisner, Chile, October 25, 1928". Ranks in appearance between an Oncilla and an Oncifelis. - A Hamburg skull no. 41996 "Felis tigrina female, ded. Hagenbeck June 25, 1927" is also published. It is still juvenile; I am putting it here because of the lack of Pm 2. - Skull Munich State Collection, 1916, No. 38, "Mrs. Ing. Schultze, La Paz (Bolivia), Chungamoyotal, 2000 m, Ihuagati". Also immature. This specimen, which most likely belongs here, comes from a Bolivian high Andean valley.
Allen's (1919) illustrations of the skull of a male harmonize with his view that the shape of the Noctifelis skull is closest to that of Oncilla. But the specimens mentioned here do not confirm this. Some of them are very similar to that of an Oncifelis, others intermediate between Oncifelis and Oncilla. In contrast to this, there are more prominent anterior corners on the zygomatic arch, broader and more expansive, which is also clearly visible on the head of a living animal Fig. 11 Plate IV. Again, the size of the bulla fluctuates. Pm 2 is absent from all. The overall impression is most likely that of a reduced oncifelis skull, conspicuous in the underdeveloped crests, in the disappearance of the weak Pm 2, individually in their somewhat slimmer structure that leans towards the oncilla form.
2). Pelt material, a) normal colors (Fig. 12a and b, panel V). - Stuffed piece from the Munich State Collection "Felis guigna Molina, Valdivia, Leybold, male". Typical colouring and spotting, as are the following, unless otherwise noted; whole length 67/68 cm, tail length 21/22 cm. - Hamburg Museum: No. 40350, same number as the skull. Whole length about 65 cm, length of tail 15 cm, male; No. 42274. Whole length 63 cm, tail length 11 cm, female. - Berlin Museum, skins or pelts: Collection "Dr. L. Plate, Lago Llanquihue (Chile)", which lies south of Valdivia, in the highlands between the main and coastal cordillera. No. 21250. Whole length 73, tail length 24 (25) cm; No. 21252. Full length 65, tail length 22 (23) cm; No. 21256. Whole length 70, tail length 19 cm; No. 21258. Whole length 62, tail length 17 cm. - Also: No. 7212 "Around Jujuy, P. Neumann". Not quite grown up. Whole length 58, tail length 18 cm. Jujuy is the capital of the northern province of the same name in Argentina, on the river of the same name. This flows to the Parana. So the piece already comes from the eastern fall of the Andes. No. 7444 "Noctifelis, male juv., Uruguay, Z. G. Hannover". No more than juv .; "Uruguay" cannot have been the home country, only the reference country. Whole length about 71 (difficult to measure), tail length 23/24 cm, in any case long tail. Turin Museum: No. 1581 "Felis guigna Molina (?), Chili, dai Missionari Francescani, 1898" Reddish brown tone, very small, juvenile piece. The presence of jagged and hook-shaped spots make this fur appear aberrant. In any case, it is significant in terms of development dynamics that in this species is not very modifiable in terms of coat pattern, there is also a variation in spotting similar to that in related genera.
b) Melanotic pelts. Melanism has been demonstrated in several genera of South American small cats. I previously knew of it as a rarity in the Margay, a species that frequents the same area as normal coloured, rosetted Oncilla pardinoides pardinoides (Gray) (Munich material from Barbieux, Matto Grosso) and from Berlin specimens of the Noctifelis guigna (Molina), one of which came from a site with normally marked specimens. The phenomenon was already known in the species (Allen 1919). You can usually still recognize the type of spotting on melanotic small cats and then determine the genus.
Pelt 21262 "Lago Llianquihue, Chile, Dr. L. Plate". Reddish black-brown. Can be safely distinguished from the pelts of small melanotic oncillas by the density of the spots, their small size, and the lack of any suggestion of rosetting. Whole length 71, tail length 21 (22). No. 21264 "Noctifelis, Chile, Chairman Albert". Somewhat softer and deeper pile. Tail torn off, but recognizable as short tailed by its thickness. Body length 53 (54) cm. Markings and colouring the same as the previous one.
Allen (1919) depicts a distinctly long-tailed male and a distinctly short-tailed female specimen and gives measurements of the total lengths and tail lengths of 7 specimenss, all from "Temulco, Chile". The largest of them has a total length of 70 cm, so that the 80 cm specimen I have measured appears exceptionally large, at least expanding the scope for this average "smallest" South American cat. Allen does not comment on the importance of the difference in tail length. His measurements of 4 males and 3 females originating from the same area resulted in overlaps. In my material you could clearly distinguish long and short tails, initially regardless of gender, whose extremes differed widely. The short tails are not only relatively thicker, they aret absolutely thicker. There is no difference in the number of dark rings. Both types fluctuate between 12 and 14 along the length with a spot-like fragment at the tip. The Berlin skins consistently have up to 13-14 stripes without any difference in length. Tail lengths as a proportion of overall lengths: 3.3; 4.3; 2.9; 3.0; 3.6; 3.6; 3.6; 3.4. The wide range between the extreme lengths is clearly evident here. The gender of our Munich specimens can be determined with certainty. It is a male with a relatively small tail length, 3.2 of the total length, less than one third. The animal described and depicted by Antonius who, according to his information, was also a tomcat, on the other hand, presented itself as a decidedly long-tailed animal. Likewise, of the Hamburg specimens mentioned above, the male is long-tailed and the female is short-tailed.
According to this, in the context of this information, the obvious differences in tail lengths that are a characteristic of Noctifelis can hardly be attributed to sexual dimorphism. Nor do they appear to be locally determined, and everything speaks against age differences. With regard to the possibility of a sexual difference, more extensive material will have to be examined, because there could be a particularly wide overlapping range of lengths with different upper and lower limits for the sexes. Nothing more can be said about this at the moment. After species distinctions based on tail lengths have been established for other genera of South American small cats - Margay macroura (Wied) vs Margay tigrina wiedii (Schinz) according to Allen (1919) - investigations into the meaning of such differences will have to be extended to these.
All previous authors named the immediate and wider surroundings of Valdivia (Molina, Poeppig, Philippi, Allen) as the area where the Guigna occurs, individual places cited were those of the coast. Molina knows the species "from the woods", Philippi found it "very often in the province of Valdivia" My material indicates a much wider distribution area: horizontally north and east, vertically up into the highlands of the Andes. There it was also found on the Ostabfall, in the Argentine northern province of Jujuy, at about 23 degrees south latitude, which drains to Parana. As a mountain dweller, it inhabits both slopes, just like the Montifelis mihi colocolo (Molina), which is close to the pointed-eared lynchailurus and continues to be traded here. The skull mentioned as being from the Bolivian northern province of La Paz, from Jhuagati, Chungamoyotal, from an altitude of about 2000 m, is very likely to be placed here as well. The relatively low elevation of around 2000 m, compared to more than 3000 m for the city of La Paz, suggests the eastern slope of the Andes for this occurrence as well. The northern limit of the distribution would then shift to between 20 and 14 degrees. Proof of substantial expansion in the opposite direction, again on the east side of the chain, was the aforementioned juvenile specimen from the "hinterland of Deseado (Patagonia), Dr. Konrad Kayser, Erfurt 1933", which obviously belonged to the species. From the eastern slope of the Chilean-Patagonian stretch of the Andes, from which the Deseado River flows, to its confluence with the Atlantic Ocean near the coastal town of the same name, the distance hardly exceeds that from the Rhine to Passau. According to this evidence, whether the specimen came from the mountain slope itself or from a place along that route, the north-south area of the species extended from the 23rd (or even the 14th to the 20th) to about 47 degrees southern latitude, without there being any significant change according to the horizontally greatly different climates or the conditions at such different altitudes. There is also information from the vicinity of Valdivia, from which the terra typica Guigna was known as "very common", which proves that it not only occurs frequently in the coastal area (according to Philippi 1873), but also in the mountains (Lago Llanquihue ). Melanism, in some of this material, is not limited to the mountains, because Philippi already calls it "very common" on the coast.
[Terra typica ľ area where the biological type specimen originates.]
Even if Allen (1919) is right in stating that this is a particularly clearly differentiated species among the South American small cats, it appears, especially with regard to the structure of the skull (see above), a generic separation of this strictly small and solidly spotted cat from Oncifelis is hardly justified. Since I do not want to make a decision on this based on my material, I have retained the special generic name Noctifelis here. Compared with the species gracilis mihi, which I (uncertainly) placed in Oncifelis, guigna agrees more with Oncifelis geoffroyi, which is typical for this genus.
Regarding the Guignaĺs lifestyle, I have already mentioned Antoniusĺs remarks about his zoo specimen. Philippi (1873) says: "It often comes to the courtyards to steal chickens, and sneaks into houses to steal meat. Some years it appears in droves, and I was told that once more than 20 of them were killed in one morning in the large meat scraping" [sic!] "In the city of Valdivia. The animal easily becomes trusting and tame." Philippi contrasts guigna with "F. Pajeros which remains uncommonly wild when captive, an impression I cannot confirm after I observed the London specimen of the same type (fig. 15 plate IV). In any case, according to this, the Guigna also appears interesting as a group, forming, to some extent, sociable "cultural followers". Perhaps it was a mountain dweller from home and, obeying its nature, only later reached the lively coastal areas. Its current relationships with small and large settlements requires surveys.
Lynchailurus Severtzow
Lynchailurus (subgenus) Severtzow, 1. c., Typus Felis pajeros Desmarest, Nouv. Dict. Hist. Nat. VI, 1816, based on the Pampas Cat, F. Azara, Essais Hist. Nat. Quadrupedes of the Province of Paraguay.
Pajeros J. E. Gray, Proc. Zool. Soc., London 1867.
Lynchailurus Allen, Rep. Princeton Univ. Exped. to Patagonia, III. part., I (1905).
Dendrailurus Pocock, Ann. Mag. Nat. Hist., 8th Ser., Vol. 20 (1917), partim (for Lynchailurus Severtzow).
Lynchailurus Allen, 1919, 1. c.
In Severtzow 1. c. Subgenus of Felis. Allen (1919) adequately justified the invalidity of the genus Dendrailurus Severtzows, to which Pocock wanted to assign the genus Lynchailurus by the same author in 1917: F. strigillata, or the putative F. colocolo Molina as described by Hamilton Smith, whereupon J. A. Wagner (Schreber's Mammals, Suppl. II) used that name, are unidentifiable. The genus name Severtzows, based on strigillata, is thus obsolete. The detailed analysis of this can be found in Allen 1. c. P. 372/73.
I stated at the beginning of this work the extent to which the genus Lynchailurus contrasts with all of the "round-eared" rosetted and spotted South American small cats. Likewise, the fact that sharply dividing differences in the skull, which correspond to this contrast as well as that of the facial structure have not yet been found (here Fig. 5, Plate II, Fig. 15, Plate IV and Fig. 16, p. 30), while those of Oncifelis and Lynchailurus appear especially similar to one another. The comparison to the "round-eared" spotted cats also applies, as far as it can be inferred from the knowledge of furs, for the new subgenus Pseudolynx to be described here and for the new genus Montifelis I am establishing for Felis colocolo Molina, which is close to Lynchailurus.
The shape of the contracting pupil was observed by O. Antonius (1. c.) In L. pajeros braccatus. He finds it "broad, if not entirely circular"; and my London specimen (fig. 15 plate IV) of L. pajeros pajeros, gazing directly into the light, shows it to be approximately oval. These representatives are thus in contrast to the old-world Felis, which they resemble in overall form. For the other lynchailurus, the pupil shape must be determined.
I. Subgenus Lynchailurus Severtzow
Corresponds totally to the list of subspecies in Allen (1919) with the addition of a new subspecies that I have found. One of the characteristics is the presence of a "mane on the back", emphasized by that author, which stands out most clearly in the rough-haired forms (Fig. 5, plate II), in the woolly-haired forms it increasingly disappears into the fur. I believe I must group this subgenus in two ways, assigning part of the "subspecies" to each. One, for which the species name pajeros is reserved, contains the Eastern South American forms with a previously known overall distribution from Matto grosso southwards to southern Patagonia; a statement in H. Burmeister (Descr. phys. Republ. Argentine, Vol. III, 1879), which extends the occurrence of a form belonging here to Chile, can refer to the other species, which, by its markings, includes western South American small cats (Ecuador, Peru) (garleppi, thomasi).
1. Lynchailurus (Lynchailurus) pajeros (Desmarest)
With blurred markings, sloping towards the back, made up of similar, usually separate spots, which in the case of the long-haired forms may be limited to the abdomen and legs, and even more suppressed. According to their way of life, Gray (1867, I.e.) counts these representatives among the "diurnal cats". The eye and face indicate that they stay in open areas ("Pampas cats").
a) Lynchailurus (Lynch.) pajeros pajeros (Desm.) amended F. Schwangart
Le Chat Pampa, F. Azara, Essais sur lĺHist. Nat. Quadrupeds Province of Paraquay, Paris 1801.
Felis pajeros Desmarest, 1. c., 1816 a. Mammlogie or Descr. des Especes de Mammifferes, Paris 1820.
Felis pajeros, Waterhouse, Zool. Voy. Beagle, Vol. II (Mamm.), 1839.
Felis pajeros, dĺOrbigny et Gervais, 1. c., 1843 and 1849.
Felis pajeros crucina, Thomas, Ann. Mag. Nat. Hist., Ser. Vol. 8 (1901).
Felis pajeros crucina, Allen, 1. c., 1905 and 1919. Nec. Allen 1919. Lynchailurus pajeros pajeros Desm.!
The descriptions of Azara and his successors do not refer, as Thomas and Allen thought, to the shorter-haired, rough-coated ones of the non-"black-footed" forms, but to the longer-haired, soft-coated ones that Thomas supposedly called crucina. Azara: "Very long fur over the whole body"; all bands of markings, with the exception of those on the legs, throat and stomach "are so barely perceptible that it might be better not to mention them "; similarly with Desmarest. Compare my fig. 13 plate V. Regarding areas of occurrence, these authors call it "south of Buenos Aires" (35 and 36 degrees). To the north it just misses Azara in Paraguay. The later information from Thomas and Allen know of locations much further south and agree with Waterhouse, to whose material from Santa Cruz (Patagonia) Thomas referred when he separated the southern form. Allen states Rio Gailegos, Patagonia, 51 degrees for this. "Felis pajero Azarae" in Burmeister (1. c., 1879) should be considered a collective description with regard to the individual forms. Since he thinks that Waterhouse's picture exaggerates the length of the fur, he may have used the short-haired form as the master (see below L. pajeros braccatus). Its location information relates to areas of various forms: entire province of Buenos Aires, more rarely Entre Rio; as already mentioned, "Chile" probably refers to one of the western genus relatives, none of which have yet been mentioned. DĺOrbigny and Gervais also refer in my opinion to the actual shape: "Colour is not very yellow, the stripes on the sides have almost disappeared, deep black leg rings." One specimen stands out with its almost pure grey and particularly long fur. The illustration shows a little more yellow, but it otherwise fits here. Gervais (Voyage de la Bonite) described the dentition of one specimen, Blainville, according to those authors, described the skeleton. d'Orbigny (1847) names 35-40 degrees and Patagonia, especially on the banks of the Rio Negro, as a distribution zone, so a suitable one for this form. Likewise, F. pajeros Desm of Schreber-J. A. Wagner (1921) also here: "Corresponds in size to our wildcat, but has a squat shape, smaller head, shorter tail and much longer fur" (?) etc.
My material: Skins from the Munich State Collection: Otto Becker 1927, No. 395 "Bahia del Fondo, Territorio Santa Cruz, Patagonia". Received on September 20th, 1927. Whole length 94 cm, tail length 25 cm (with end hair 26 cm). The coloured areas of fur are a pronounced silver, woolly. Back mane almost level with the thick fur. Covered in spots as far as abdomen, excluding the legs. The whole of underside, the hind legs, and the front legs down to the paws have large, strong spots. The base colour on the underside from the chin to the end of the abdomen is milky white, as are the undersides of the forelegs and hind legs. The upper four large spots on the forelegs form perfect bands around the leg. There are streaky spots on the thighs as well. Rear soles are slightly reddish. Relatively short, thick, bushy tail, in stark contrast to the long, thin tails of the northern forms. Ears smaller than those (a consequence of its distribution, according to Allen).
Otto Becker 1930, No. 137 "Comodoro Rivadavia (Chubut), Patagonia, 13. 1. 30, total." This summer coat is essentially the same as the previous September coat, only the base colour is a little more yellow, it is less woolly and the guard hairs stand out a little better. Waterhouse (1. c.) of his two pieces, one from April, one from August, unanimously: "Most remarquable the length of the fur." The markings, on the other hand, change "greatly in intensity" with the seasons, which does not apply to our specimens.
Skin from the Turin Museum: "Felis, Patagonia". Long, shaggy, woolly hair, similar to Munich specimen No. 395, but with a slightly more yellowish tinge. Stripes on the side of the trunk indicated. Marking features similar, pronounced rings on the legs. Soles without markings or streaks. Tail (torn off) thick, woolly. Ears with strong tufts, whereas those in good condition show only a hint in the form of thicker, longer hair. I remind you of the individual occurrence of ear tufts in domestic cats as in their wild ancestors F. ocreata Gm. Still a juvenile.
Skulls that should be assigned to this form have not been presented to me. Allen's skull picture (1905) was not accessible to me. Its dimensions (1919) reveal a broad, apparently quite massive skull. O. Thomas finds in his case the "only difference" compared to the northern form is a "noticeably larger width of the posterior palatal opening", but correctly adds that this particular dimension is inconsistent in the felids. The description of Blainville's skeleton (see above) should be referred to when comparing felid skeletons.
According to the information available, including my own, the previously secured distribution of this form extends from about 35 degrees to 51 degrees south latitude, which means that it almost reaches the southern tip of the continent. From this point of view, the Berlin piece No. 9216, that will be mentioned, represents a type of transition to the eastern tropical forms; we also find transitional copies of the other type, as will be shown. According to Azara, his long-haired Pampas cat, which he knew from northern areas of the distribution zone, "prefers the cold and temperate regions" there (hence he associated them with the lynx). So perhaps in latitudes where these occur alongside the northern forms there is an overlap according to climatically different habitats.
Divergent skins: Hamburger Museum No. 38537 "Felis pajeros Desm. $. Zool. Garten, June 8th, Brazil "(Fig. 14, plate V). Extremely woolly, similar to the fur of a coyote, even surpassing Montifelis colocolo in this regard. Coloured areas, with eight ninths of the back hair length being yellow-red, with light gray tips, awn hairss in between with fox-red tips. With the exception of those on the tail and (suppressed) on the front legs, there are no marking features, either on the underside or rear legs, which makes the piece stand out from the previous ones. A dirty light yellowish grey predominates below and on the legs. The thick, woolly tail is blurred near the base, but still clearly ringed with black-brown, even having complete rings, seven in total, and the tip is black-brown. Ears not standing out in colour, thick tufts of hair standing out from them. The top of the head bears small, blackish spots down to the nose, which is reminiscent of the Asian Manul (Otocolobus manul Pallas). The mane of the back is lost in the hair, its region being powdered with black like the Munich pelts. Total length 87 cm, tail length 30 cm. Despite the statement "Brazil" in this zoo specimen, it must either be a far more southerly or a more pronounced mountain form, possibly both, which, judging by this one piece, almost gives the impression of species diversity. I believe I am justified in assigning it the rank of a subspecies special form, whereby it remains to be determined what causes the differences, which can certainly not be considered seasonal, I choose the name
b) Lynchailurus (Lynch.) pajeros hyperthrix n. ssp.
The original subject of my photo of a living lynchailurus (Fig. 15, Plate IV) from the London Zoo, where I saw the animal in 1931, should also be included here. In terms of its coat properties, it corresponds perfectly to the description above. Differences: The almost complete disappearance of the markings also on the tail, its somewhat more prominent emergence on the front legs, a still distinct darkening on the ears, the fading of the blackish dots on the head. The broad, compact shape of the head and face will already be developed or initiated with pajeros pajeros. It corresponds to the above-mentioned picture of Allen's skull. - Type specimen of this Subspecies is the aforementioned fur No. 38537 of the Hamburg Museum.
The second of the divergent skins, which is close to L. pajeros pajeros, is of interest as intermediate between it and the rough-haired form L pajeros braccatus, the description of which follows here. Berlin Collection No. 9216 "female, Lyncailurus, Tornquist, Argentina, Zool. Garten, d. July 13, 1897". Still immature. The density of the fur is intermediate between that of L. pajeros pajeros and that of L. pajeros braccatus. The basic tone of the coloring is closer to grey than to yellow. In the emergence of markings again intermediate between the two mentioned; the intensity of the leg markings puts it closer to the Munich pajeros pajeros.
c) Lynchailurus (Lynch.) pajeros braccatus (Cope) emend. F. Schwangart
Felis braccata, Cope, Amer. Nat. Vol. 23 (1889).
Lynchailurus pajeros braccatus (Cope), J. A. Allen, 1. c., 1919.
Lynchailurus pajeros pajeros, J. A. Allen, 1. c., 1919, nec. Desmarest 1816!
Felis (Mungofelis) braccata (Cope), O. Antonius, Der Zool Garten Vol. 6 (1933).
Allen mistakenly understood Desmarests and his successor L. pajeros to be a rough-haired, relatively short-haired form, while, as shown above, the original descriptions referred to the later named "crucina" by Thomas, to which the name pajeros must be transferred as a subspecies. Gray changed the name to Felis pampanus (1. c., 1867) for unknown reasons, describing a skull. It is possible that this description was based on the form in question here; then this would have to bear the well-chosen name pampanus. Our knowledge of the skull formation in the individual lynchailurus is insufficient to make a decision. Anyway, I have reservations. Sclater (List of Vert. Anim., 1872) changed pajeros to "passerum", according to Allen only to eliminate the "barbaric pajeros". Cope described his Felis braccata as a special species due to its deep black-brown fore and hind feet, which distinguish it from its closest relatives. Everyone leaves it undecided whether it is a melanotic characteristic or a local form ("pajeros braccatus"). His representation of the Copes type does not reveal the black coloring of the feet. Since our material contains transitional pieces, while geographical separation of the final phases is out-dated, I believe that I cannot maintain the subspecies separation from the previous basic form. O. Antonius (1933) even suggested the establishment of a special subgenus Mungofelis for braccata Cope; the peculiarities in posture and behaviour, which he describes in his specimen in the Schoenbrunn Zoo, are certainly in common with other lynchailurs, just as his illustration (fig. 5 plate II) as well as the excellent head study by H. Krieg (Fig. 16) should be considered typical for the basic form in terms of art-physiognomy. Both contrast the typical L. pajeros pajeros in the same way as the L. pajeros hyperthrix n. Sbsp. (Fig. 15, plate IV). - Since the previous designations of the basic form were either definitely incorrectly related to this, or at least probably incorrect (Gray 1867, Sclater 1872;) it only remains, if one does not want to separate braccata, to relate their names on the whole, bearing in mind my previous reservations.
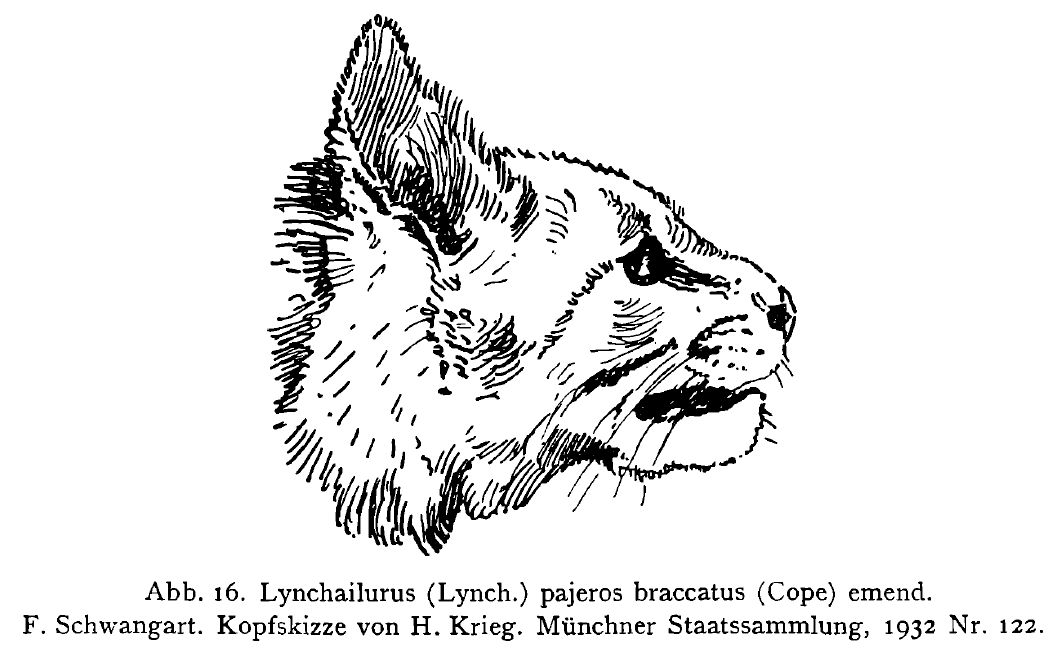
Fig. 16. Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Head sketch by H. Krieg. Munich State Collection, 1932 No. 122.
Members of this overall type form the stock of the subgenus Lynchailurus in the tropical steppe area of eastern South America. In type, these cats differ from those of the two previous forms (p. Pajeros and hyperthrix) by their shorter, thinner, rough, straw-like hair without "undercoat", the pronounced brownish, often fawn-brown background tone of the coloured regions of fur on the upper side, the yellow to yellow-brown on the underside, on which deep brown blurred spots stand out, with the exception of an only very short whitish chin zone, together with the corresponding somewhat stronger markings of the brownish backgrounded thighs and especially due to these features: clearly contrasting, long-haired, dark mane, very long, thin, loosely haired tail (Figs. 17-19, plate VI, skins; Fig. 5, plate II, living animal, compared with Fig. 15, plate IV). The complete pictures show differences in the width and length of the head and face.
The parallel between the conformation of Lynchailurus (Lynch.) Pajeros and that of the ancient genus Felis (in the narrower sense, without Catolynx chaus) is evident here. In both cases we are dealing with a geocircle [ring-species/cline] whose representatives living in warmer climates are straw-haired, relatively large-eared and long-tailed, while those in colder climates are shaggy to woolly-haired, relatively small-eared, short and thick-tailed. This contrast corresponds to "AllenĹs rule". I have given more detailed information on this in my monograph on domestic cats (1932, p. 14); there also something about the naming of such geographically determined clines and their relation to the concept of species. The usual naming of such forms as geographical or ecological "races" is given by a further term, breed, as a name for inherited, selectable mutations of every characteristic, according to which in the wild there are also melanotic representatives of species, which are also common in the small South American cats, which if the trait were inherited - which we do not yet know - should be called "races". I have proposed "geocircle" [ring-species/cline] and "geotype" [landrace] as unequivocally narrower, limiting expressions for the case of geographically determined shapes and forms and, against the criticism (in the zoological report) which found this suggestion superfluous, I hold to my view that this specification is objectively justified. - With Felis, the "cold forms" inhabit the north, the warm forms the south of the total area, with the pajeros it is necessarilyy the other way round.
If in the geocircle pajeros one looks at the braccatus form in the terms just explained, one tries to expand the parallel with Felis even further, especially in the area of the latter form. Within braccatus one notices the blackening of the feet in some specimens, which, as already mentioned, has led to attempts at species or subspecies separation (Cope, Allen). With Felis, the southern forms (Ocreata) have a dark stripe on their back feet, while the northern forms (Silvestris) only have a short spot in its place. Since braccatus, in general, occupy a tropical to highly subtropical area, the difference, if at all geographically, could be more vertical. In the meantime, we do not have the necessary data for dissemination; much more study material is required. Allen suspects a "regional" form with black feet. More on this under phase C. Here I will introduce a transitional form of braccatus in terms of foot colouring; we do not know of such forms in Felis. Even the northernmost ocreata form, F. ocreata sarda, has the dark sole stripe.
My material:
Phase A (without black feet). Fig. 17. Plate VI
Hamburg Museum. Skins No. 42804 and No. 42805 "Felis pajeros Desm., Dr. Schreiber, N. Jenny, Paraguay ", No. 23276" F. pajeros Desm. var., female, Zool. G. 23. 4. 1897 ". In the first two, the pattern appears unevenly distinctive, which is important for assessing its modifiability given it is from the same location and date. The third coat is a little more woolly, the hair thicker, also stronger and longer on the inside of the ear. All three are still young. Skull No. 23277 for the third coat is exceptionally wide in the scale of this form, with maximally protruding front zygomatic angles that appear triangular. The breadth of the zygomatic arch and palate are particularly noticeable in this still young and feminine piece. In a second Hamburg skull, No. 23811, "Pajeros, Argentina, female", these characteristics are less developed. It comes from a relatively small but adult animal.
Paris Museum. Two pelts "F. Pajeros, Wagner, Argentina, 1911". One lighter with a grey tinge, the other with a foxy background color.
Phase B (intermediate stage for blackening of the feet). Plate VI. Fig. 18
Munich State Collection. "Barbieux, 1931, No. 81, cat", location is missing. Whole length 111 cm, tail 39 cm (pulled). Weak, longitudinal row spotting found on the sides of the trunk, only blurred due to a slight color difference. On the soles, the hairs of which have been poorly preserved, darkening isindicated along the length, but no actual sole stripe. - "H. Krieg, expedition 1931/32, bought in Assuncion." Length of trunk 65 cm, tail end torn off. Base distinctly tawny coloured, almost like the "Jaguarondi" (phase of Herpailurus), reddish yellow on the underside; slightly shaggier hair than typical for the subsp. Spotting on upper parts almost suppressed by the deeper brown of the background. The front and rear feet have chocolate-colored markings up to the joint, specifically a sole stripe, which does not yet take up the entire surrounding area, but is interspersed with a background colour like that on the trunk.
These two Munich skins, Barbieux 1931, no. 81, and H. Krieg 1931/32, from Assuncion, are thus two intermediate stages between the form without black feet to the black-footed form (CopeĹs/ AllenĹs "braccata"). From other articles by Barbieux we can infer this specimen was from the "Matto Grosso" area.
H. Burmeister (1879) had apparently also seen a specimen with transitional colouration of the foot: "Sole of the foot almost pure black", found in the area of Entre Rios province, northern Argentina.
Phase C (black-footed). Fig. 19 Plate VI (fur), Fig. 16 in the text on p. 30 (head).
Munich State Collection. Exp. H. Krieg 1931/32, No. 122, male, "St. Luis, Apabergland ". Skin, skull and skeleton. Total length of the skin 81 cm, tail length 28 cm. Probably still immature. Basic color yellow-brown, clear row spotting with a tendency to chain formation in places. Colour distribution in no way different from that of typical specimens of the previous phases, with the exception of the black-brown fore and hind feet up to the joint. The proportions of the skull resemble that of a slender member of the genus Oncifelis. The zygomatic width, the width of the nose and the palate, the distance between the canines, etc. are all relatively considerably smaller than in the Hamburg skull No. 23277 belonging to phase A (see above), where that was a female, here it is a (probably younger) male. Allen (1919) presents a skull from phase C (specifically his "braccatus"), and gives its dimensions, narrow compared to one from phase A (his "pajeros pajeros"). This is also that of a female. We can rule out this difference between the skulls of the phases as a sexual characteristic, since in all these small cats, as expected, the male skulls tend to be broader.
The few previously known samples of this phase occupy the zone of the total distribution of Lynchailurus pajeros which is closest to the north of the equator. According to Allen, CopesĹ type specimen examined by him comes from Matto Grosso, i.e. below 15 degrees south latitude. According to O. Antonius, the Schoenbrunn piece was "bought from Bolivian traders"; the buyer, a Mr. Hofmann, saw the animal "very often in the highlands of Bolivia" (although it remains to be seen whether this observer could always have paid attention to the colour of the feet). H. KriegĹs very typical piece comes from the Apabergland, west of the Rio Paraguay, below about 23 degrees south latitude, Northeast Paraguay. The mountainous country consists of "low plateaus with small mountain ranges of medium mountainous character". The samples known so far came from the hill country and low mountain range of the zone which represents the northernmost part of the distribution area of the total pajeros species (the geocircle) and closest to the equator. It remains uncertain whether phase C of Lynchailurus pajeros braccatus is absent from the associated plains or, depending on the nature of the terrain, is replaced by the other two phases, i.e. whether it fully deserves the name "mountain cat" attributed to it by Antonius.
From the description of O. Antonius (1932) about the behaviour of his zoo fosterling (Fig. 5, Plate II) one can infer peculiarities of lifestyle that are certainly also common to the other conspecifics: "Obviously it's an outright ground animal; the guard only once saw it on the resting tree, which was set low above the ground. It does not move in the manner of the supple jaguarondi "(Herpailurus, weasel cats)", but rather it walks relatively quickly on its very fine, small-pawed legs, almost dancing. . ." On the lifestyle of this group see also the introductory words on Lynchailurus (Lynch.) pajeros (Desmarest) on p. 26.
2. Lynchailurus (Lynchailurus) garleppi (Matschie)
Felis (Lynchailurus) pajeros garleppi Matschie, meeting minutes. Ges. Naturf. Friends of Berlin, 1912.
Felis pajeros garleppi Loennberg, Arkiv foer Zool., Vol. 8 (1913).
Felis pajeros thomasi, Loennberg, same place.
Lynchailurus pajeros garleppi, Allen, 1919, 1. c.
Lynchailurus pajeros thomasi, Allen, 1919, 1. c.
According to the data available so far, the Lynchailurus, which live mainly in areas west of the Andes and their region itself, form a group that clearly differs from the forms in the southwest South American geocircle, while in spite of their strong geographical proximity to those forms in some places, transitional phases are missing. Accordingly, a species-identifying naming method appears justified compared to the sub-specific ones of the authors.
A separation of the species into two forms of a lower category is no longer possible due to the additional material that I provide here. The differences in the spotting characteristics are within wide limits, but are transitionally connected. Typical representatives were previously known from Ecuador and Peru, whereas no decision is needed regarding the species belonging to the Chilean "pajeros" mentioned by Philippi (1870, 1873) and Burmeister (1879). In itself, this group is also likely to occur in Chile. The coloured pictures of "Felis pajeros Desm." by D. G. Elliot (1883), one of which O. Thomas relates to the Patagonian form and the other to the more northern of the eastern South American forms, both belong here ("garleppi"). My material provides evidence of the distribution of this species over the Andes to their eastern descent, as is the case with other mountain cats; compare here under Noctifelis guigna and Montifelis colocolo. The unmistakable closeness of Lynchailurus garleppi to an occurrence of L. pajeros braccatus is attested by O. Antoniusĺs credible source for high Bolivian territory (see above).
The species is already characterized by the sharply contrasting, distinctly longitudinally oriented yellow-red pattern on a silvery-deep grey background, which progresses in individuals from relatively small spots to chain formation with the appearance of sometimes very large central areas. In the multiplicity of these stages up to the formation of longitudinal chains with elongated central areas, this speciesĹ markings behave convergently with the large ocelots (Leopardus), where the same process is found in individuals. A second parallel phenomenon in the round-eared South American small cats is the light spot on the auricle (shown in Loennberg, 1. c.), however, this differs in position and shape from that of the large ocelots. This faint spot is easily lost on preserved skins, as was the case with my material. E. Loennberg also used its absence to characterize "garleppi" versus "thomasi". A back mane, a characteristic of the subgenus Lynchailurus, is present throughout.
My material: Berlin Museum. Skin No. 21244. The type specimen of Felis garleppi Matschie, Fig. 20 Plate VII. "Cuzco, Peru, O. Garlepp." Fairly long straw-like hair, the smallest spots of this study material (which Loennberg used in 1913 to differentiate his thomasi). Despite the tendency towards this in places, nowhere are there chain arrangements, some spots show centrl areas. Banding on the legs similar to L. pajeros. Back mane accentuated by dark colouring. Whole length 79, tail length 25 cm (probably a bit torn off at the tip). Cuzco is located in the mountainous region of the Andes; pateaus, diverse climates.
Berlin Museum. Pelt No. 1213. "Lynchailurus. Route to Bolivia over the Punta near Jujuy." Basic color somewhat lighter grey, laterally more yellow (cream) predominates. Spots paler (faded?). Spots consistently much larger, here and there central areas formed; from the beginning of the rear half of the trunk they close up into short chains. A little larger than the previous specimen. Head is missing. Tail probably a bit shorter. The location is northern Argentina, already belonging to the eastern flank of the Andes.
Munich State Collection. Stuffed piece, catalog no. 26/30. "Felis colocolo Molina, Bolivia, Coll. Printessin Therese. Bought from Indians at the market in La Paz (Bolivia), Oct. 2, 1898, poorly stuffed." The somewhat faded colouring was no doubt similar to that of the Berlin type. From the front torso, behind the shoulders, large spots in the longitudinal rows usually meet and often merge, forming chains. Almost certainly recognizable as female.
In the collection there is a skull that, according to the slip of paper attached to it, should belong to this specimen, but it is too small in comparison with it. The occipital region is missing. It is not possible to judge the development of the crest. According to the other common characteristics (zygomatic width, front corners on the zygomatic arch) it belongs to an Oncifelis or a Lynchailurus. The rear edge of the hard palate has no indentation, is simply rounded, which also occurs as an exception in South Americans (Oncifelis) and is recorded here.
Turin Museum. No. 2213. "Felis andina Thomas (?), male, Forests of the western Andean slope. NW de Quito (Ecuador)." Fig. 21 plate IV. Of a somewhat woolly character. Long parallel spots with a tendency to form chains, in places there are chains. The particularly extensive occurrence of central areas in the large areas is striking. The light spot on the back of the auricle, which Loennberg claims for the species, is clearly visible here. Closely related to the Munich piece in terms of chain and central area formation, but also goes beyond it. Even the stripes on the back are only solid in the front part. In his otherwise detailed description of a F. thomasi, Loennberg does not mention the formation of central areas, but these are visible in his illustration.
A skull "Leipold, Chile" from the Munich collection, from a relatively small adult animal, could belong to this group. But a decision is not possible.
In the current system, a criterion of species separation in cases like that of the pajeros and garleppi groups of lynchailurus is the occurrence of both in the same location, whereas in the case of local separation they are called "races", or in my terms as "geotypes" of a "Geocircle." One usually recognizes this characteristic for mere "race" ranking or "subspecies" quality even in cases of very considerable differences between the forms in question, with a certain dislike of previously prevailing decisions between "valid species" and lower categories based on the degree of diversity. According to this, some would be inclined to add Lynchailurus (Lynch.) Garleppi (Matschie) as a further "race" to this group, despite its significant differences from the forms that are united here as Lynchailurus (Lynch.) Pajeros (Desmarest). I take the position that both criteria apply and must be weighed against each other, as long as geographic transition types are not found - which is not the case here - and I think it is right to regard pajeros and garleppi as different from one another on the basis of previous knowledge.
After an inspection of the type specimen of Felis pajeros garleppi Matschie, Loennberg determined the close association of this specimen with his Felis pajeros thomasi, but also highlighted some distinguishing features. With the exception of the previously discussed white ear marking, which he found "obsolete" on garleppi, all of these features appeared transitional. The illustration of thomasi (Loennberg 1913/14, 1. c.) shows, compared with the Matschies type specimen (fig. 20 plate VII), the essential difference is the characteristic pattern: thomasi tends to form extensive central areas and short chains. Neither forms chains as much as the Turin specimen in fig. 21 plate IV, hence "thomasi" is intermediate between this and garleppi. Our Munich specimen is then intermediate between thomasi the Turin specimen.
As long as geographical contrasts do not correspond to appearance, which is linked by transitional forms, these can only be viewed as the end phases of individual variability, something that certainly also applies to the small differences in the skull of garleppi vs thomasi mentioned by Loennberg. Nevertheless, as I have said, considering the few known specimens of the type illustrated by Loennberg, this series does not represent an end phase. I would like to keep the name of the "Thomasi Phase" as the counterpart to the "Garleppi Phase", which has distinctive central areas and a clear tendency to form chains, because Loennberg demonstrated this difference.
The "Phase" concept indicates contrasts of an individual nature, regardless of whether they are modifications or alternatively inheritied traits, or the existence of any intermediate stages. The experimental determination of the inheritance of such phases is an important task in itself.
Examples linked by transitional states: the opposites "Eyra" and "Jaguarondi" in Herpailurus, the formation of rows and chains of spots in Leopardus, probably the same and the formation of central areas in the spots (rosettes) in Lynchailurus. Examples without transitional states: Melanism in the South American genera Margay, Oncilla, Noctifelis.
I do not include the phase name as the lowest category in the designation of the form, as with subspecies, geotype or ecotype. So, in contrast to those terms, it is written with a capital letter: Garleppi, Thomasi.
To distinguish between the different variants of the species Oncifelis geoffroyi, the term "Form" was used here (pp. 16 and 17); argentea form, flava form, salinarum form. I prefer this to the term "phase" if there are sufficient indications that the differences correspond to geographical or ecological separation, however, this is not yet certain or the circumstances have not yet been clarified to such an extent that one could speak of eco- or geotypes. The fact that, according to the current state of knowledge, the boundaries between phase, form and geo- or ecotype can be uncertain, means the designation is often left to the "discretion" of the systematist, and only corresponds to the nature and purpose of our research.
II. Subgenus Pseudolynx nov.
Munich State Collection; 1934, skins nos. 51-54, 58 and 59, bought from the Reiss-Munich company in November 1934. Place of purchase "Buenos Aires". The subgenus is based monotypically on the new species Lynchailurus (Pseudolynx) kriegi n. Subg described below. n. sp.
To determine a specimen as belonging to the Lynchailurus genus, I base it on the fur character, the basic colour and also the, albeit peculiar, pattern type, but especially on the pointed, tufted ear, which fortunately is fully preserved (one-sided) in skin No. 54 prepared for trade. It can also be seen in a second skin.
The shape of the pattern is my deciding factor in assigning the species mentioned to a particular subgenus. A longitudinal direction is maintained on the top of the head with many lines (forehead and top of the head), on the several stripes on the neck and is adhered to in the form of the darker, backgrounded longitudinal line along the back; there are also diffuse longitudinal spots from the nape to the base of the tail. Otherwise, the trunk is clearly patterned all over, not unlike that of a spotted tabby house cat, different from the pattern type of other Lynchailurus. The slightly thicker haired, darker longitudinal stretch of the back possibly represents the "mane" of others. The name Pseudolynx (pseudolynx) refers to the pointed, tufted ear.
Lynchailurus (Pseudolynx) kriegi n. Subg. n. sp.
Cotypes of the above-mentioned skins no.51, 52, 54 and 59.Fig. 22 a, b, c, plate VII.
The fur character is quite long-haired, very soft, slightly woolly and loose. The basic colour is a light, attractive creamy yellow which becomes gradually lighter towards the belly, but without becoming white. Basal part of the hair is grey. The entire pattern is deep brown, standing out strongly. These characteristics are the same for all skins.
The spots are individual dots of very unequal size, also very different in number, more or less stretched, while in places (fur no. 51) there is almost a horizontal striping. The transverse direction of these components of the pattern can be very pronounced or can also be somewhat disordered. Towards the underside, the spotting becomes sparse. Pronounced stripes on the hind legs. With the exception of fur no. 52, which is therefore a co-type specimen, the feet have been removed during preparation. This fur still has the left hind foot. This shows a hazy grey "sole stripe". On the tail there are half-rings only around the distal half. The distance between the rings is uneven. Ears lack any marking paler than the background colour on the back, above in the basic tone of the coat. Ear-tuft greyish-yellow.
Lengths: No. 51. Whole length 76 cm, tail length about 22 cm.
No. 52. Whole length about 84 cm, tail length about 27 cm.
No. 53. Whole length and tail length? Badly mutilated.
No. 54. Whole length 89 cm, tail end missing.
No. 58. Whole length about 78 cm, tail length 23/24 cm (end is missing).
No. 59. Whole length 87 cm, tail length 27 cm.
With regard to their home range, it should first be noted that the almost long-haired, warm fur excludes them originating from the dispatch location, Buenos Aires. It is likely to be a more southern (Patagonian?) form or it comes from a mountain region. The dealer's later attempt to determine more precisely failed. There is no doubt that the collection comes from a rather narrow find area.
Genus Montifelis nov.
Felis Molina, 1. c. 1782 (partim), species colocolo.
Felis Edw. Griffith, Animal Kingdom, Vol. 2. London 1827, spec. "F. colocolo of Molina "?
Dendrailurus, R. J. Pocock, Ann. Nat. Hist., Ser. 8, vol. 20 (1917) (nec Severtzow 1858). Invalid.
Oncifelis, J. A. Allen, 1919, 2. L. c. (partim).
Allen (1. c.) Tries to prove that the cat mentioned and depicted by Griffith in no way corresponds to colocolo Molinas and that it is indefinable. I have also mentioned the author because, contrary to the description that it is actually not identifiable and the location information does not match the colocolo, the poor image may still refer to this species, which is the only one of the new genus known to date. Pocock, who evidently did not know any original of this cat, places it together with pajeros, genus Lynchailurus, which he wrongly calls Dendrailurus, and even doubts whether the two can be distinguished. Allen (1. c.) places it with geoffroyi in genus Oncifelis, though it is "obviously different" from that species. In fact, due to its pointed ears and pelt pattern, it is closer to Lynchailurus than to the round-eared, spotted Oncifelis. However, it has characteristics that also justify its generic independence with respect to Lynchailurus (Fig. 23, Plate VI). Above all, no other cat can be compared to it in tail formation; this is extremely long and evenly thick like a cylinder from beginning to end. The marking pattern also stands alone in its extremely large spots and its grouping of spots, as well as in the overall colour, which, in combination with the thick woolly fur, makes this handsome animal probably the most beautiful South American cat. The subdued hue of all the tones represented on the fur looks particularly attractive. This animal, previously a rarity in our collections, can therefore be addressed as living in open terrain. Its fur quality suggests a mountain dweller.
What brings the genus closer to lynchailurus is bove all, as I have said, the pointed shape of the ear which distinguishes both of these from other South American cats. This shape could be seen in the skins that I used as templates, but without an actual point. Although the edge of the ear was somewhat abraded in all of these, a perfectly pointed ear is unlikely to occur even in an intact state. This is shown in the illustration, based on a living specimen in the Santiago Zoo (Chile), made by Philippi (1870), who gave the first detailed description. With regard to the reliability of this illustration, however, the author himself makes the proviso that the animal could only be drawn uncertainly due to its unrestrained nature, which we can confirm based on the very schematic representation of the drawing, and he did not provide a depiction after the animalĺs death. The stature is probably reproduced correctly in the main. It also makes the animal look strange. A "back mane", as in Lynchailurus in the narrower term (subgenus), is not present in Montifelis (colocolo).
According to the undoubtedly good skull pictures of Philippi (1873), as the text notes, distinguishing features against Lynchailurus ("pajeros", more likely garleppi) are also present on the skull. The Colocolo has a fairly long, tapering snout, which is also shown well in the portrait, and unreinforced anterior zygomatic corners. Seen from the side, the incisors protrude conspicuously. Ihering (1. c.) draws attention to the angled, downwardly curved chin, which also indicates "pajeros". However, this characteristic varies within several genera, for example in Lynchailurus themselves.
Allen (1919, 2) considered the typical habitat of the colocolo on the basis of the oldest description (i.e. Philippi) that precisely identifies the species: the Cordillera near Santiago (Chile). A type specimen for this description is not known, but it is probable enough that material which should be considered typical is available in Santiago. Philippi drew his full image based on a specimen in the local zoo; he does not state whether the animal was preserved after death. He also fails to state whether the two images of a skull (from the side and from above) that he delivered three years later, were from the same animal or even which specimen they come from. The other pieces to be mentioned here complement the distribution picture.
The genus is based monotypically on the species colocolo, as can be seen from the following.
Montifelis (gen. N.) Colocolo (Molina)
Felis colocolo, Molina, Saggio sulla storia nat. del Chili, 1782.
Felis colocolo, Philippi, Arch. Naturg. Year 36, Vol. 1 (1970). - Derselbe, Felis colocolo, Arch. F. Naturg., Volume 39, Vol. 1 (1873).
Felis colocolo, H. Burmeister, Descr. Phys. Republ. Argentine, 3rd vol. (1879).
Felis jacobita, Cornallia, Mem. Soc. Ital. Sc. Nat., 1865.
Felis jacobita Com., Matschie, meeting minutes. Naturf. Friends of Berlin, 1912 (as "race").
Felis (Lynchailurus) colocolo neumayeri, Matschie, 1. c.
Oncifelis colocolo (Molina), J. A. Allen, 1919, 2. 1. c.
Oncifelis colocolo neumayeri (Matschie), Allen 1919, 2. 1. c.
A large, handsome cat with thick woolly, clearly contrasting, but modestly tinted fur bearing very large markings and a very long, evenly thick tail cylindrical from its base to its blunt tip, encircled by around 10 rings, including one at the very end that is still unbroken. On the feet there are broken ring formations split into spots. The background colour is creamy yellow, which is slightly smoky on the top, which is mainly concentrated on the head, ears, neck and shoulder region, with a sharp boundary against the back behind it. On this background, from the back of the head to the base of the tail, there is a beautiful reddish-brown to orange-yellow marking which is arranged in a striped, ornamental manner in the region with the darker base colour. Further back, along the largest part of the trunk, there are rows of spots, the rows essentially running sideways even if the spots appear to form diagonal and longitudinally arranged markings. (Philippi's picture, 1870, shows stripes to a greater extent than in all the skins that I examined.) The back of the ear does not have a light spot, as is found in all round-eared genera and among garleppiĹs lynchailurus, the ear-tip is not tufted.
My material: Munich State Collection; 1925, no. 257, bought by H. Krieg from the dealer Tutzer, Buenos Aires, "Mountains of Tucuman or Catamarca". Total length 113 cm, tail length 47 cm (Fig. 23 Plate VI). Pattern and colouring as just described. The coalescence of spots here and there simulate markings with central areas. On the hind feet, shadowy grey respesents a "sole stripe."
Berlin Museum. "No. 34507, Lynchailurus, La Riocha, Argentina. Tutzer ges., Zukowsky ded." Total length 111 cm, tail length +45 cm. Colour contrasts are a little stronger than the previous one. In the rear part of the median of the back, the red-brown spots form a kind of stripe where they are strung together.
Berlin Museum. "No. 34508." Same Denomination of Origin. Total length 114 cm, tail length 46 cm. Almost the same as the previous one. Mr. Zukowsky had correctly mentioned these two Berlin skins as "colocolo" in a letter.
Milan Museum. The type specimen of Felis jacobita Cornallia, which Allen (1919) recognized as being identical to colocolo Molina. I examined the fur in Milan. The stuffed specimen is called "Felis jacobita Cornallia, F. pajeros Desm. var.? Argentinia, donated by Dr. P. Mantegazza ". There is a skull in it, but given its small size, it is improbable that it belongs to the fur. Size of a domestic cat. The fur is essentially the same as the one in Munich and the two in Berlin. Back and side pattern is less prominent in this one; some of the large side markings can be chaaracterised as rings.
Berlin Museum. Type specimen of "Lynchailurus colocolo neumayeri" Matschie. "No. 34506, Rio das Mortes, Matto grosso, Dr. Neumayer ". As a precaution, Allen (1919) still lists this specimen as a subspecies. The supposed "differences of colouration" do not exist; it resembles the Munich copy in the background colour and hair pattern. Details, such as the fact that in places the spots merge into transverse bands "less distinctly" (based on the aforementioned PhilippiĹs figure), turn out to be modifiable characteristics evenwhen considering my small amount of study material.
Occurrence: Philippi's specimen came from the Chilean Cordillera, near Santiago. It was caught at a Hacienda Dehesa near a location called "Infernillo". Burmeister (1879), who saw it in Buenos Aires, added to the Milanese "Felis jacobita" specimen that it had been shot in the mountains near Potosi or Humacuaca in Bolivia. Potosi lies in the middle of the southern part of the Bolivian Andes branch; the province of the same name extends across plateaus to the west of it to the Chilean Andes. The mountains rise up to 6500 m. Tucuman and Catamarca, which Tutzer stated as the region of origin of the Munich sample, are in northwestern Argentina; the province of Catamarca in the eastern mountain range of the Argentine Andes rises to 6000 m. La Riocha, where the first-mentioned Berlin specimens are from, is a province in Argentina, bordering the Cordillera and Chile to the west and is mostly mountainous.
According to this information, colocolo, already a mountain animal according to its general build, colonizes both slopes of the Andes and probably also their plateaus and elevations up to significant heights on Chilean, Bolivian and Argentine territory. The distribution limits are uncertain given the small amount of material.
The location mentioned for the "Lynchailurus colocolo neumayeri" (Matschie) specimen: "Rio das Mortes, Matto Grosso" does not agree with this distribution. A region called Rio das Mortes is located in the Brazilian state of Minas Geraes, which contradicts the addition "Matto Grosso". However, this area of discovery would also contradict everything that is otherwise known about the range of the species. Matto Grosso is separated from the Andes and their foothills by large areas of Bolivia. The location information is questionable from the start, despite the statement by Matschie (1912) that the piece was "shot" by the donor. In the information I requested, Prof. Dr. H. Pohle, head of the mammal department of the Berlin Museum, said with good reason that it is easy to confuse the place of discovery in this case. Due to the arrangement of all the material from that haul it was very likely that they had been bought together. The reference locations reached as far as Oran. He comes to the conclusion, with which I can only agree, that the true place of discovery is "not ascertainable."
Bibliography
(Individual forms are noted at the head of the description)
Allen, J. A. Severtzow's Classification of the Felidae. In: Bull. Am. Mus. Nat. Hist., Vol. 41, New York 1919.
Allen, J. A. Notes on the Synonymy and Nomenclature of the smaller spotted Cats of Tropical America. In: Bulletin of the Mus. Hist., Vol. 41, New York 1919.
Antonius, O.: About Felis braccata Cope and other small cats from Schoenbrunn. In: The Zool. Garten, N. F., Vol. 6, Leipzig 1933.
Bemmelen, J. F. van: Colour pattarn of mammals. In: Verh. Deutsch. Zool. Ges., 34th annual vers. Utrecht. 5. Supplement vol. Zool. Anz., Leipzig 1931.
Birula, A .: Contributions to the classification and geographic distribution of mammals. VI. On the position of Otocolobus manul (Pallas) in the system of the Felidae family and its races. In: Annuaire Mus. Zool. Ac. Sc. 21, Petrograd 1916.
Brehm, A.: Tierleben, 2nd edition, Berlin 1876.
Haltorth, Th.: The relationship of the big cats to one another. Diss. Univ. Berlin 1937.
Henke, K.: On the morphology and developmental physiology of animal markings. In: Die Naturwiss. 21st year, Berlin 1933.
Ihering, H.: Systematics, Distribution and History of the South American Predators. In: Arch. Naturg., 76th year, vol. 1, Berlin 1910.
Kretzoi, N.: Materials for the phylogenetic classification of the aeluroids. In: X. Congres intern. Zool. Budapest, Budapest 1929.
Krieg, H.: On pigmentation in mammals. In: Anat. 54th vol., Jena 1921.
Krieg, H.: Principles of striped markings in mammals, derived from studies on solipeds. In: Lect. and up. Development mech. of organisms. Issue 30. Berlin 1922.
Krieg, H.: Stripes and movement. Attempt at a causal definition of some of the basic elements of mammalian marking. In: Arch. Development Mech. Vol. 51. Berlin 1922.
Krieg, H.: The zoological problems of the Gran Chaco. In: Zool. Number Vol. 74. Leipzig 1927.
Krieg, H.: From the Andes of Bolivia to the Atlantic. In: Ztschr. Der Ges. F. Geography of Berlin. 1938.
Krieg, H.: As a zoologist in the steppes and forests of Patagonia. Munich-Berlin 1940.
Krieg, H.: and H. Foerster: Basic rules of dorsal markings among the Anura. In: Zool. Anz., Vol. 119. Leipzig 1937.
Loennberg, E .: Notes on some Cats from Eastern Asia with description of a New Subgenus. Section: A short review of the structure of the anterior portion of the zygomatic arch in Felidae. In: Arkiv foer Zool., Vol. 18. Stockholm 1926/27.
Pocock, R.I.: On Domestic Cats. In: Proc. Zool. Soc. I. London 1907.
Pocock, R. I.: On the Hyoid Apparatus of the Lion and related species of Felidae. In: Ann. Mag. Nat. Hist., Ser. 8, Vol. 18. London 1916.
Pocock, R.I.: Some Dental and Cranial Variations in the Scotch Wild Cat (Felis silvestris). In: Ann. Mag. Nat. Hist., Ser. 8, Vol. 19. London 1916.
Pocock, R.I.: On the External Characteristics of the Felidae. In: Ann. Mag. Hist. Nat., Ser. 8, vol. 19. London 1917.
Pocock, R.I.: The Classification of Existing Felidae. In: Ann. Mag. Nat. Hist., Ser. 8, vol. 20. London 1917.
Rode, M. P .: On the Use of Selected Fur Characteristics in Mammal Systematics. In: Bulletin du Museum, Ser. 2, vol. 3. Paris 1931.
Rode, M. P.; Comparison between the pelage of Felids found by M. G. Babault in the region of Kivu (Congo Beige) with the pelage of golden cats from Africa and Asia. Study of furs. In: Bulletin du museum. Ser. 2, Vol. 4. Paris 1932.
Schneider, K. M .: Observations on the shape of the pupil in some living mammals. In: New psychol studies, 6 vol. Munich 1930.
Schwangart, F.: On Racial Constitution and Breeding of the Housecat (Results and Problems). In: Ztschr. Fur Mammalkunde, Vol. 7. Berlin 1932. Also in book form.
Schwangart, F.: The Manul, Otocolobus manul (Pallas), in Felid Systematics. In: About true cats and civets. Central Jurnal "Small Animals and Furry Animals" (Carnivore Studies II), year 12. Leipzig 1936.
Severtzow, M. N.: Notice on the multiserial classification of Carnivores, especially Felids, and the studies of general zoology which are attached to it. In: Rev. et Mag. Pure and Applied Zoology. Ser. 2, Vol. 10. Paris 1858.
Toldt, K.: Epidermal stripes, hair banding and wild markings in the house cat. In: Verh. Zool.-Bot. Ges. Wien, vol. 62. Vienna 1912.
Toldt, K.: Structure and natural colour of the coats of wild mammals. Leipzig 1935.
Thomas, O.: The spotted Tigercats of Brasil (Notes of Neotropical Mammals). In: Ann. Mag. Nat. Hist., Ser. 7, Vol. 12. London 1903.
Zarapkin, S. R .: On the question of the relationship between the big cats. In: Ztschr. F. Mammalogy, Vol. 14. Berlin 1939.
List of figures
The full body pictures of furs, Leica photos by the taxidermist E. Schuhmacher-Munchen, on which are based length measurements given in the text in natural size relationships to one another. After this and in connection with the dimensions in the text, direct details of the reductions are unnecessary, where they could not be negotiated with the producer in the field.

Possibly the same pet Margay seen by Schwangart.
TABLE I
Fig. 1: Margay sp. juv. "Dondoca". Intern. Cat show, Paris May 1931. From: Revue Feline de France, Paris 1931.
Fig. 2: Margay tigrina (Schreber), phase Mitis-tigrina mihi (which will be reported on later). Joinville, southern Brazil. Zool. Hamburg Museum. - a) male, No. 39723, b) female, No. 39729 of the museum. Phot. E. Schuhmacher-Munich.
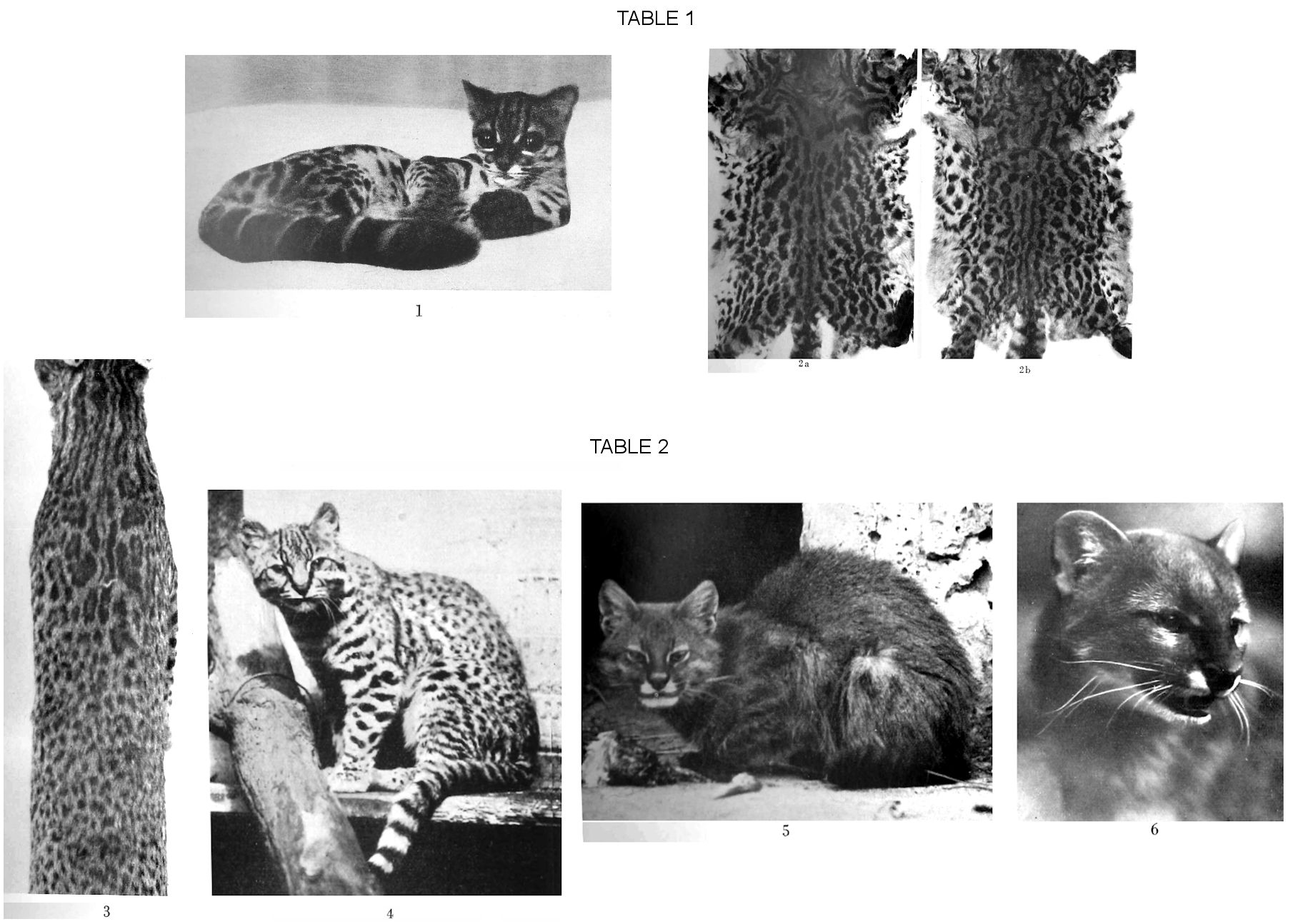
TABLE II
Fig. 3: Oncifelis geoffroyi (dĺOrb. And Gervais), forma flava m. Shoulder drawing, exceptional ring formation. Museum Berlin No. 13278. Phot. E. Schuhmacher-Munich.
Fig. 4: Oncifelis geoffroyi (dĺOrb. And Gervais). Photo of the "Association for Cat Breeding and Protection", Dresden 1928.
Fig. 5: Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Vienna Schoenbrunn Zoo. Phot. Rob. Weissenbach-Vienna.
Fig. 6: Herpailurus jaguarondi (Lacepede). Photo Elfe Schneider, Zool. Garden Berlin.
TABLE III
Fig. 7: Oncifelis geoffroyi (dĺOrb. And Gervais), forma argentea mihi. Co-type specimen, 1930, No. 136 of the Munich State Collection. Photo E. Schuhmacher-Munich.
Fig. 8: Oncifelis geoffroyi (dĺOrb. And Gervais), forma flava m. Individual transverse rows of spots on the sides of the chest. Berlin Museum, No. 13278. Phot. E. Schuhmacher-Munich.
Fig. 9. Oncifelis geoffroyi (dĺOrb. And Gervais), forma salinarum Thomas. Munich State Collection 1932, No. 48. Photo E. Schuhmacher-Munich.
Fig. 10: Oncifelis gracilis n. Sp. Munich State Collection, 1934, no. 55. holotype. Phot. E. Schuhmacher-Munich.
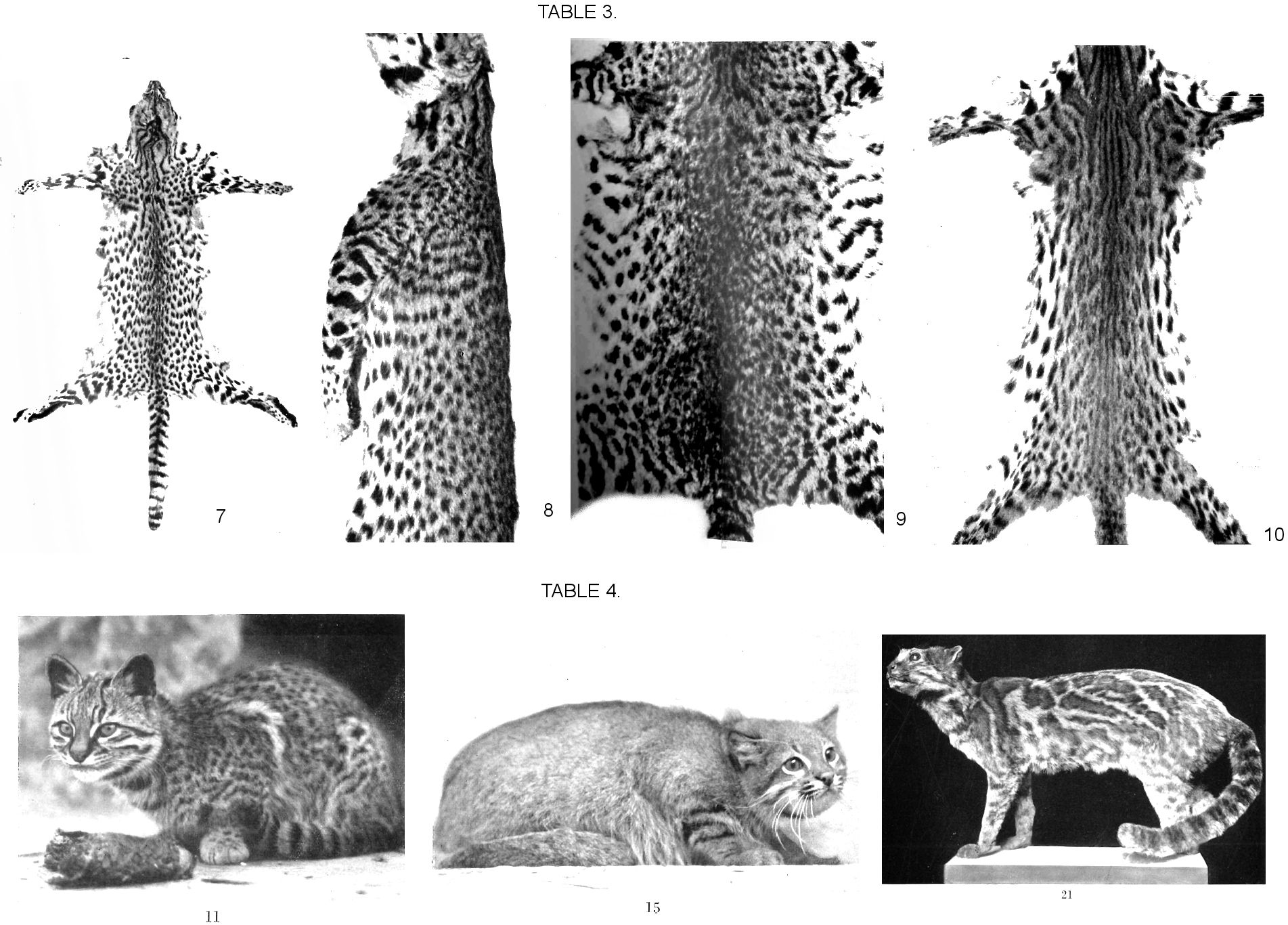
TABLE IV
Fig. 11: Noctifelis guigna (Molina). Vienna Schoenbrunn Zoo. Phot. Hanni Glassner.
Fig. 15: Lynchailurus (Lynch.) Pajeros hyperthrix n. Subsp. Zool. Garden, London.
Fig. 21: Lynchailurus (Lynch.) Garleppi (Matschie). Spots with large central areas, chain formation. Turin Museum, No. 2213. Phot. Prof. Dr. Archangeli-Turin.
TABLE V
Fig. 12: Noctifelis guigna (Molina). Museum Hamburg, a) long tail, male, no. 40350, b) short tail, female, no. 42274. Phot. E. Schuhmacher-Munich.
Fig. 13. Lynchailurus (Lynch.) Pajeros pajeros (Desm.) Emend. F. Schwangart. Munich State Collection, 1927, No. 395. Phot. E. Schuhmacher-Munich.
Fig. 14. Lynchailurus (Lynch.) Pajeros hyperthrix n. Subsp. Museum Hamburg, No. 38537. Phot. E. Schuhmacher-Munich.
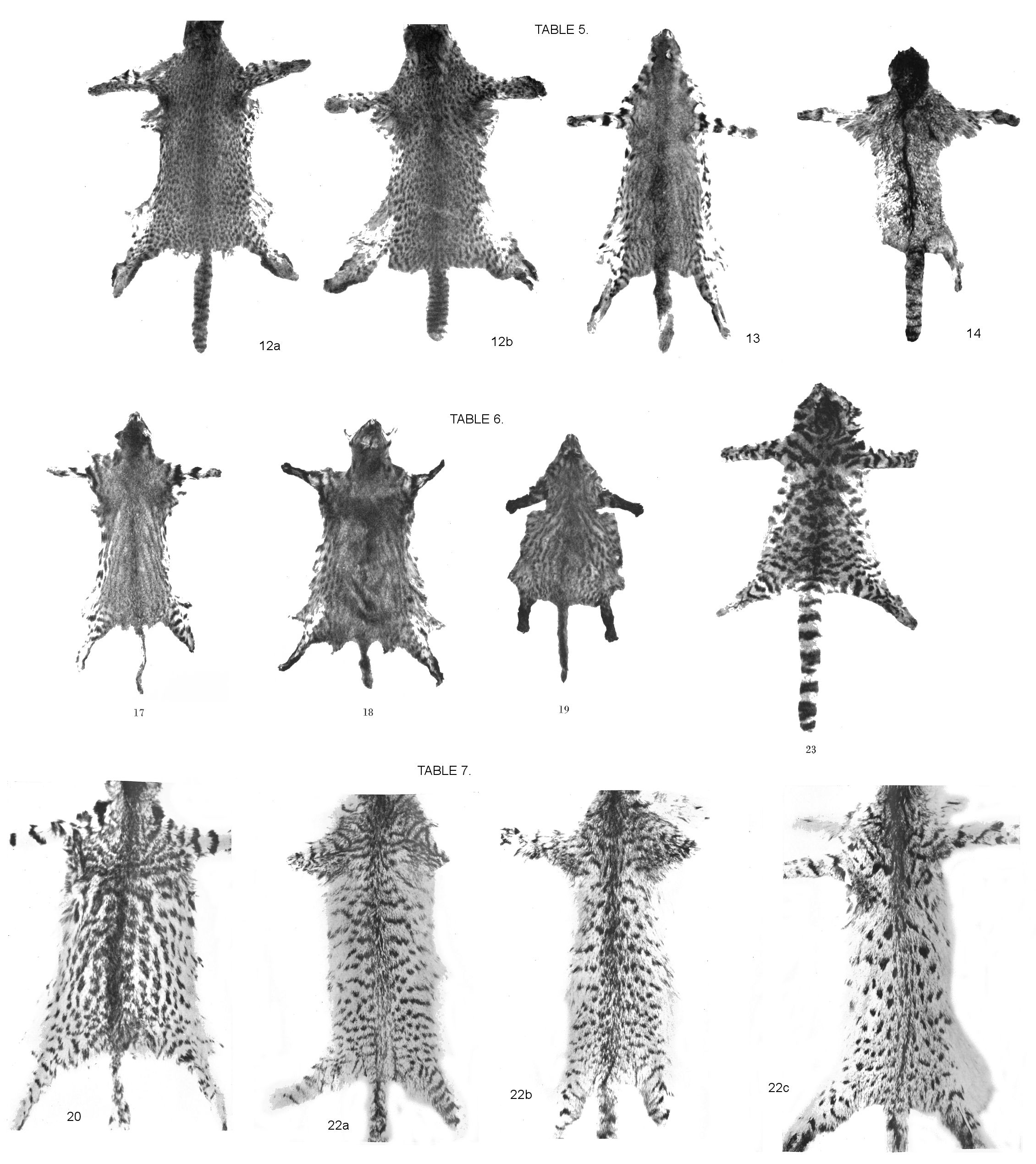
TABLE VI
Fig. 17: Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Phase A mihi. Mus. Hamburg, No. 42805. Photo E. Schuhmacher-Munich.
Fig. 18: Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Phase B mihi. Munich State Collection. Exp. H. Krieg 1931/32. Photo E. Schuhmacher-Munich.
Fig. 19: Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Phase C (Copes Pha Munich State Collection 1932, No. 122. Photo E. Schuhmacher-Munich.
Fig. 23: Montifelis n. Gen. Colocolo (Molina). Munich State Collection, 1925, no. 257. Photo E. Schuhmaker Munich.
1925, No. 257. Photo E. Schuhmacher-Munich.
TABLE VII
Fig. 20: Lynchailurus (Lynch.) Garleppi (Matschie). Type specimen. Museum Berlin, No. 21244. Phot. E. Schuhmacher-Munich.
Fig. 22: Lynchailurus (Pseudolynx) kriegi n. Subg. n. sp. Munich State Collection, a) No. 51, b) No. c) No. 54, 1934, cotypes. Phot. E. Schuhmacher-Munich.
IN THE TEXT
Fig. 16 (p. 30): Lynchailurus (Lynch.) Pajeros braccatus (Cope) amended by F. Schwangart. Head sketch H. Krieg. Munich State Collection, 1932, No. 122.
TRANSLATOR'S ADDITIONAL NOTES ON THE PET SOUTH AMERICAN SMALL CATS
From the French Journal "HIS HIGHNESS THE CAT," Country Life, and Farms and Chateaux (Vie a la Campagne), Special Issue, 15th April, 1935. Volume 92.
ôDONDOCAö, THE MARACAJA FROM BRAZIL
PEOPLE OF LETTERS have always been very interested in the Cat. This was the case of Ms. Marcelle Adam whose words, published in this magazine a few years ago, you will read with pleasure.
ôDondocaö, a Maracaja from Brazil (Maracaja, in the Indian language, means Woodland cat), is a small Tiger-cat. He was born in one of the lush forests where the deep waters of the Amazon flow. An Indian took him from the nest after having undoubtedly killed his mother.
What a wonderful animal of the equatorial jungle! His flexible, thin, elongated body wears the richly spotted dress of the panther. The strong tail, very long, hangs to the ground and acts as a pendulum during the acrobatic exercises required for hunting in the trees. Capped with round ears, always restless, the head is small, or rather it seems small, because its extraordinary eyes take up all the space.
Those immense and unfathomable eyes, velvet eyes of an almost black brown, those prodigious eyes, they have a look so soft, so uneasy, so sad, so mysterious, that it penetrates and transfixes us. We bring it with us, it haunts us and moves us. What is this little Brazilian Tiger-cat dreaming of, captive in a Parisian home where our concern vainly strives to make it forget the tropical splendours?
It seems obvious to us that this nocturnal being does not tolerate daylight well and that he is happier in the shade. Despite our care, he remains fearful. His fears, always fresh, give birth to formidable reflexes, and his narrow jaw is armed with strong teeth in the form of fleur-de-lys, like those of the big cats. We donĺt think heĺs nasty. Perhaps he would even be sociable if we managed to cure him of his atavistic anxiety. He has bonded slowly with us, and his friendship remains steadfast. His memory often surprises us. He is filled with intense caresses for Mr Pierre Orcheboeuf, who acquired him from the Indian hunter who captured him.
M. Pierre Orcheboeuf, crossing the sea, brought back this precious and turbulent friend, who still did not know how to drink or eat. Then he sold it to an American, who dared not keep it, because training it requires patience, courage and leniency! It also requires boundless dedication, because a Tiger-cat does not easily adapt to our polite manners, nor to the hygienic customs which the Emperor Vespasian first thought of.
One day, Mr. Pierre Orcheboeuf brought me ôDondocaö (that was the name of this little panther). You know that I deal with novelists and Felines with equal zeal. They both have fairly similar manners, with one difference, as our friend Paul Brulat would say, that Novelists begin by congratulating each other before scratching each other, while Felines, slow to make friends, start off by scolding and bristling, and stop tearing each other apart once they know each other.
So I adopted ôDondocaö. The Tiger-cat began to lay down the law at my place, not supporting any authority, wandering through the ornaments as through the jungle and fixing his dark, cloudy coloured eyes on us. His body is elongated, very level, supported by strong legs higher at the back than in front like those of the lynx. ôDondocaö does not meow like a Cat. He emits a kind of yelp when he is angry that reminds of the cough of his bigger cousin, the Tiger. When he is bored, he makes a kind of plaintive and prolonged yawn. When he is very happy, he sucks his thumb, absolutely like a small child. In front of his chopped meat, he growls as if defending prey.
In short, despite the incompatibility of our characters, I tried to live on good terms with ôDondocaö. I would never have asked for a divorce if ôManou de Madalpourö, a sacred cat from Burma, had not been threatened by this bloodthirsty South American. Foreseeing a possible drama, I resigned myself not to divorce, but to the separation of body and property.
ôDondocaö now lives with one of my friends. He seduced her. As she is sensitive and generous in character, he abuses her goodwill. He rules over her, he makes a martyr of her; yet she admires him with the eyes of a happy slave and finds something exotic and delicious in her torments, - Marcelle Adam.
THE TIGER CAT
Of remarkable dimensions, a hunter and jumper of the first order, it is very cuddly and affectionate. [The photo shows a mackerel tabby domestic cat.]
There existed in France, a few years ago, a curious specimen of the Tiger-cat, imported from the Argentine Republic and belonging to Mrs. F. Marie, who provides us with the following description:
This Cat, which does not seem to be the product of any special, selected breed, was born from the probable mating of a very beautiful Cat with a Wild cat from one of the northern provinces of the Argentine Republic. This four-year-old Tiger Cat weighs 8 kg. about. Despite this respectable weight, its short coat gives it a rather slender appearance. Unlike the appearance of Cats, which have a short, rounded muzzle, its muzzle is longer, somewhat pointed, and slightly hollowed out under the cheekbones. Very small, his nose has a funny profile, seeming to start from the top of the skull. His head is finely marked with very clear black lines which contrast with the very bright yellow-sepia background colour. His eyes, half closed during the day, are very open at night.
Standing very tall on his legs and also quite long in the body, he is perfectly proportioned. His legs are very sinewy and have a trigger; he can spring, without momentum and without effort, a height of 2.5 m. He never tires of repeating this leap, much to the admiration of observers. His gait, in step, is quite characteristic: he has a way of ôrollingö his shoulder blades that is very reminiscent of the look of the wildcat; when he runs, he looks like a galloping racehorse. He has a low, horizontal tail, flush with the ground.
His agility is such that often, when seeing a bird on a branch distant from the trunk and 3 m. from the ground, instead of trying to climb the trunk, he crawls on the ground and leaps onto his victim, whom he very rarely misses. He hunts with passion. His natural wildness appears on occasions and is sated on backyard birds in particular. He brings in the results of his hunt with satisfaction and even pride, and the admonishments he receives do not quench his thirst for blood.
Note - the accompany image showed Dondoca the Margay and two "Tiger Cats" (one of which is a tabby domestic or hybrid, the other a wild cat).
