ON RACIAL CONSTITUTION AND BREEDING OF THE HOUSECAT (RESULTS AND PROBLEMS)
By F. SCHWANGART (Dresden).
With 27 illustrations in the text and on the panels VI-XV.
MAIN SECTIONS:
I. Summary of Contents (with page name)
II. Introduction, Position of Cat Breeding
III. Basic Breed Types and Characteristics, Historical Tribes
IV. The Typical Design
V. Colouring (of hair, skin, eyes etc.)
VI. My Shorthair Breeds and Their Structure
VII. The Overall System
VIII. Mental and Physiological Factors. Merits and Breeding Lines
IX. Conclusion, Bibliography, Clarification Tables
TRANSLATOR’S NOTE.
Some ofthe terminology used by Schwangart does not translate directly into modern English.
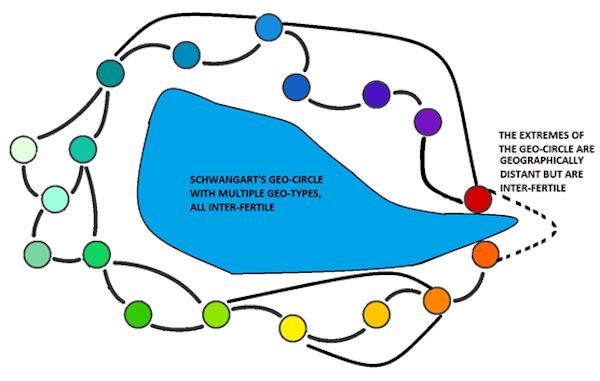
A Geo-circle, or Geo-group, is a group of closely related sub-species with localised differences (unlike a ring-species, the extreme ends are inter-fertile). A Geo-type is a regional type. Modern equivalents might be races and sub-races. Type-circles and Form-circles are used in the same way.
The term “Rassen” has different meaning depending on context: race, species, sub-species or breed.
The term “Angora” is generally used to mean “Longhair” while “Half-Angora” means “Semi-longhair.” At one point Schwangart mentions “Angoras” from the place of that name, meaning “longhaired cats from Angora.”
I. SUMMARY OF CONTENTS.
(Keywords are highlighted in the [original] text of the essay by bold or spaced type.)
Pg. 76, The move towards efficient breeding. The current situation.
pg. 77, Elements of physical appearance: shape, colouring and markings. The character principles of long hair (Angora) and its three breeds. - The Persian. Biological basics of its head shape. Its relationship to colour. The German Longhair. My reasons for creating it. Analysis of its characteristics. Obstacles in breeding practice.
pg. 81, Main features of Longhairs and the phylogenic history of the tribe. F. silvestris SCHREB. as an ancestor. F. ocreata GM., Ornata GRAY and silvestris SCHREB. a geo-circle. Angora cats in Angora.
pg. 81, Relationships within the Geo-circles and from there to the domestic cat. The Manul's alleged claim to ancestry. F. silvestris and ocreata as extremes. Continental and Mediterranean contact, differences in the closer Ocreata grouping, the Balearic cat. The Geo-circle [species family] and Bergmann's and Allen's Rule, Indications of Phenotypic Induction and principles for this. The question of fertile hybrids between the Geo-circles, and the impact on housecats with respect to parallelism. Colour and markings. F. Ornata‘s relationship to F. silvestris, F. ocreata and the long-haired housecat. Area and grouping of the closer Ornata family.
pg. 89, “Half-Angoras,” heredity experiments and practice, notable special cases. The Birman breed: description, origin and reservations. The Siamese: the “Annamese breed”. Breed-creation by crossing different cat breeds.
pg. 92, Contrasts in terms of the scoring of physical characteristics in the Longhair and Shorthair in hitherto English-led breeding. The cult of colour, creation of a range, suppression of different types, and disharmonies. Assets of this method of breeding.
pg. 93, Cases of scarce older form utilization in domestic Shorthairs: 1.)
pg.93, The blue-short hair: The preference for “blue”. The unsatisfactory types, “Russian” and “British blues”. The tendency of the Blue Shorthair to further changes in conformation; excessive conformation [extreme type]. Comparison with the Blue Longhair. “Carthusians” [Chartreux] (“Maltese” - 2.)
pg. 95, The “Abyssinian” (“Nubian”). It’s relationship to wildcat type. Its similarity of colouring and change of conformation. Degradation (sudden change). Its nature and the nature of the Nubian pattern. The question of racial worthiness.
pg. 98, The common colour patterns. Special remarks about the Tiger pattern. Status in the three wild groups. Development History Data and Hypotheses. Phylogenetics, systematics and importance of defining breed. Breed standards and practicalities. Degrees of weakening compared to the different domestic colours, colour and pattern combinations. Reduction and reinforcement of patterns and colours in domestication. Modalities [types/phenotypes]. Exaggeration of stripes, transitional characteristics. Intensification of infantile traits. Types of repression. Asymmetries. Prospects of racial hygiene [breed purity] value of the patterns. Scientific inheritance.
pg. 101, From the marble [classic tabby] pattern: Isolation of the domestic cat by its two-tone pattern. Is this pattern a feature of the type or is it a domesticated mutant? Testimony on the depth of domestication. Equalities and differences in the behaviour of the patterns. Construction, variability and “basic types” of the marble pattern. Mixing and transition between the two patterns. Interpretation of both cases. Assumptions about origin, and trigger causing the marble pattern. Question of blends, possibility of re-establishing the tiger pattern inside the marble pattern. Is it influenced by environmental factors? Changes during postembryonic life. Previous experiments crossing the patterns. Considerations of their constitutional or eugenic significance. Pure breeding of the marble pattern. Favourable circumstances, difficulties, guidelines.
pg. 109, Importance of markings in the domestic cat as a basis for research into the causal laws of animal markings (including the state of research).
pg. 111, Regarding the hair colouring of the domestic cat:
pg. 111, The wild colour. Generality, artificialness, changeability, constitutional priority. Comparison between wild colour, wild pattern, and marble pattern. Silver gray and slate gray. Causes of the constitutional primacy of wild colours and relatives. Counterpart in humans. Review of the marking patterns. Comparison with the similar colour and marking differences of animals in the wild state, also with regard to the constitutional significance.
pg. 114, Black. Melanism in the wild state. Sequence from white to black and vice versa. Rustiness. Persistent foxy black brown. Gap with “red”. Two kinds of black? Counterpart piece in humans. Divided opinions about melanism. Status with the cat. Pronounced inclination with black to slenderness. Shorthair and long hair. Acro-melanism and related traits. Black and the marble pattern.
pg. 115, White. Leucism and albinism. Acro-leucism and acro-albinism, both in a broader sense. Special type of white marking in the “smoke cat”. Propagation tendency of white; Steps, process, with piebald whites in general, with the acro-leucism (acro-albinism) of the domestic cat and in the wild state. Breeding acceptable whites. General judgments regarding pathological characteristics of albinism and leucism. Full and partial albinos in the house cat. General constitutional weaknesses in whites? The particular defect deafness linked to blue-eyed white, looser link to any white markings. My breeding conclusions. Question of the importance of subtle differences in the partially albino cat's eye. Long and short hair. - Inheritance experiments with white spotting.
pg. 3 20, Flavism. Views about its constitution. Status and peculiarities of “Siamese dilution”. Relationship of Flavism to white (especially to albinotic) and to the patterned “reds.” General constitutional weaknesses and special deficiencies in the Siamese. Interaction of various causes.- “Red dilution” in heredity experiments. Special case of siamese dilution.
pg. 122, Blue. In the wild F. ocreata. Review of its relationship to conformation. “Relaxation”, but weakness of the constitution has not been proven. Breeding rule for “blue skin”. Difficulties of maintaining purity. Steel blue and silver blue. Inclination towards whiteness over the silver colour. Blue Tiger, blue marble. No direct transition to black and white? Opposite of the diluted “red”; as well from blue to red in the position of break between blue and black. Blue and white piebald. Incorporation of black for brightening up. Contradiction between practice and attempt and means of its mediation. Blue and the “Annamese breed.”
pg. 123, Overview of the experimental results of cross-breeding characteristics of the various colour groups. Special remarks about “tortoiseshell” and “Spanish”.
pg. 125, Privileged position of the domestic cat regarding the investigation of domesticated influences. These are still very much at work here. Inbreeding and weakening, neglect, warmth and cold, pregnancy confinement and suckling.
pg. 126, Coat colours. Importance of researching colours to determine the rules. Investigation. Results and conclusions. Practical academic problems. Skin colouration in nude cats. Desirable amendments. The oral cavity and ball of the foot.
pg. 129, Different colour of the nose leather. Crimson as a characteristic of F. silvestris (according to E. SCHWARZ) and in domestic cats. Investigation. Results, conclusions, problems.
pg. 130, Eye colour. Its relationships, questions. Colour of the claws (and foot pads).
pg. 131, Importance of body shapes in connection with pattern and colour of Domestic Shorthair. My racial divisions and the thereby exploited form differences. Occurrence and attention even in ancient Egypt. Hints of F. ocreata material. Perfection on the slender side by the ancient elite after the change caused by crossing it to F. silvestris. Inheritance of some characteristics together. Absence of hereditary studies. Breedability of the two family groups. Individual reasons for this. Ratio of body size to the family grpoups. Ratio of both to the standard limits.
pg. 135, Expansion of breed forms. Additional details: nose curve, ear shape, forehead, nose profile, hair character. Apparent and actual secondary impairment of the shape in the case of thinner animals. Types of combination of each of the two family groups with colouring or markings in the “Shorthair” and “Slim” breeds. Pros and cons of these types of combination. Position of the smoke, isabelline and cream-coated cats. The classic tabby [marble] Shorthair.
pg. 138, Constitution of the three Shorthair breeds. Restricting this breed basis to shorthair. Standard of breeding in the various breeds of cats and the respective methods offered.
pg. 139, The shorthair stock excluded from my three Shorthair breeds and its fate. Breeding classes without breed. Reasoning. The “Belgian”.
pg. 140, The basic features of my whole breeding classification and their expression in the “Standard”. Individual principles. Comparison with officially used classifications.
pg. 142, Physiological and psychological basis for breed creation. The performance point of view also with the domestic cat. The cat as a working animal, and culturally significant pet. Corresponding properties. Abilities, character. Research and utilization. Its usefulness as a reason for utility breeding. Mouse-exterminator. Short- and Long-hair. Fitness as a ratter and selection for performance, systems for doing so. Types of testing and evaluation. Additiona pet control: weasels, ermine, polecat, vole. Selection possibilities.
pg. 146, Domestic cat and fur-farming. Fur animals (selection). Direct Utilization: illicit mass occurrence, worthlessness for fur farming. Conflict between pet and fur animal evaluation.
pg. 147, Favourable variations of instinct. Carrying of prey, proper retrieving, accompanying, leashes, types of attachment. Selection and and training. Correct abilities. Longhair and Shorthair. Preferences and difficulties.
pg. 148, Unfavourable variations of instinct and the question of their breeding direction. “Spraying” tomcats, overlong or imperfectly retracting claws, “bird addiction”.
pg. 150, The various breeding principles outlined here and their practical relationship to each other. Significance of the results. Presentation of progress, among other things, at a survey of the treatment of the domestic cat in major modern animal breeding faciilities.
II. INTRODUCTION, POSITION OF CAT BREEDING.
The purpose of this work is the announcement of a critical principles and also a generally breed-based significant approach to cat breeding and the data that made this approach possible. The main reason of general interest comes in a discussion of my “breeding classification and assessment rule” O. Antonius (1931 c), with reference to the special feature of the case, “show the emergence of breed types in a house-pet so far almost completely free of artificial breeding.” This applies mostly to the so-called “ordinary” Shorthair, often just called the “domestic cat,” where breeding for type is a custom not followed here. But it also applies to the long hair, which has long been specially bred for its form, and where we find ourselves in the extremely instructive position of having before us a plastic material that is still open to the creative and reformatory influence of scientifically based, stringent selective breeding. It is already expressed in these last words that it is not only phylogenetic and inherited scientific hints that guide us in establishing breeders of valid breed types, guiding the selection and combination of traits, according to which “raciality”, as I would like to call it, decides on an overall appearance.
In a recent work by SCHWANGART and GRAU “About deformity, especially hereditary tail deformities, in the domestic cat” (1931), I dealt with the importance of scientific breeding, and also considered the aesthetic, ethical and utility factors. The value of performance should be considered in such an immensely useful, indeed indispensable, pet as the cat. In the meantime we considered only the prevention of misbreeding and the required abilities that characterize performance, but specifying attributes for specialised uses seems problematic. I will return to the consideration of performance, as well as considering the mental and physical qualities that make up the pet's virtues, with ethical and aesthetic considerations in mind. Compared to the psychological variability of the dog, the cat – which has not be developed into types - seems to me to be greater in different directions. Almost nothing has happened to it from a selection point of view. The heritability of such variations is often found.
I dealt with tribal history, interrelations and heredity data in my work published in 1928 and I provided an additional version in my booklet of 1929. In this book I will consider the various questions that were not sufficiently discussed previously. Since those publications, there have been valuable contributions to our knowledge.
The breeding biology situation, as noted in the passage quoted earlier in a review by ANTONIUS, gives the cat scientist practical and scientifical trump cards in the hand. Usefully, by the detailed presentation of breeding lines, at the same time it obliges him to caution in relation to primitive stock untainted by aesthetics; ugly cats are exceptions among the uncultivated stock. My scientific decision to publish my book proves that my preoccupation with these topics is fruitful, but I must ask the reader to take into account the drawbacks of the stuation, particularly with regard to the use of details. It is based on an unusual backwardness of most cat owners who feel like breeding cats (including “leading personalities”), compared to the level of other contemporary animal breeding, and on the awful addition of our ambivalence towards the native Shorthair, which is the result of the vilification of cats through one-sided bird protection propaganda. The slump is favoured by German cat books of a type that owes its existence to an insignificant luxury boom, to teach them about nature and pure breeding, they resemble more uncritically illustrated collections of legends and easily present a seemingly absurd standpoint, the slowdown of an economic situation would be more beneficial to the continued existence of a branch of breeding. Another strong barrier is the lack of a reputable professional journal with similar objectives as the new “Journal of Dog Research”.
III. Basic Breed Types and Characteristics, Historical Tribes.
From the appearance of the body, the cat breed makes use of three basic elements: shape, colouring, and markings. To breed utility forms and varieties little challenges feline zoology. In my work with Dr. med. GRAY on “Deformity”, I took a different position. There are plenty of breeding form variants available, but they are mostly overlooked by the up to now more playful breeding direction. But it likes to pursue blatant deformities of pathological-degenerative characteristics (such as those tail deformities) and seeks out such disfigured animals to produce “breeds”. I want to bring it back from this principle. At least, I would apply this to dwarf cats as soon as this feature could be firmly shown to be inherited in animals.
Decades ago, English cat breeders decided on a single form of long-haired cat, the “Persian”, which they have recognized since a breeder convention of 1887 (fig. 4). Since then, it has rejected a second type of Longhair known as “Angora”, because such exhibition animals were bred to be either identical with good Persians or they were seen as inferior. The same can still be stated, where so-called “Angoras” occur in cat shows. I use “Angora” as a collective name for all longhaired cats, in the sense of the term “Angorism” when this condition occurs in any kind of short-haired breeding animals. “Angora” is thus synonymous with “long hair”, and the English Persian is a Longhair or Angora.
Other long-haired breeds are the German Longhair (Fig. 5), which I have created myself, and the Birman breed, which has been exhibited in Paris for two years (Fig. 6). With regard to conformation, these common features apply first to the Persian and German Longhair:
“Firm body, short, stocky legs, broad head, with relatively short, wide-ended muzzle. The ears moderately small. Fairly short, beautifully carried tail. Pronounced, soft long hair (allowing for age differences, season, possibly pregnancy). Back parting, 'frill', 'breeches' on the hindquarters.” (SCHWANGART “Breed Classification and Evaluation Regulations”.)
I must emphasize after this diagnostic analysis the inadmissibility of slender form, narrow head, Spitz-like sharp muzzle in the German Longhair as well as in the Persian. For there is the erroneous opinion that these were always defects in the Persian and were admissible, or even essential, in German Longhairs so that one could now pass off flawed animals, formerly known as “Angoras”, as “German Longhairs.”
Especially in Persian, as opposed to German Longhair, these features are required:
“Thick, round head, protruding forehead, abruptly dropping away to the broad, short bridge of the nose, forming a saddle as they meet, which together give a 'grimacing' expression. Hair covering is a bit woolier than other long hairs. Breed for size and strength. Good development of 'frill' desirable.”
Already in my treatise on “Deformity”, etc. (1931) I drew attention to the Persian head having the domestication feature of “Short Face”, but it is not allowed to reach the degree seen in dog breeds, where the upper row of teeth recedes behind the lower; this deformity (see ANTONIUS 1922, pg. 57) in the true “Pug” being accompanied by regression of the chewing muscles. This foreshortening produces the round head of Pugs and similar breeds. Let’s examine the state of these muscular parts as well as the “round head” of the Persian - there is a lesser degree of muzzle-shortening, but the change in the forehead and skullcap can be said to have progressed in the same manner as the Pug.
On the whole, the typical Persian head seems to me to have been caused by two components. Once one recognizes as a basis the head and face form of the Nordic type of the wild Felis silvestris SCHREB. (Fig. 7). Added to this are the domesticated deviations just described. I regard this second group of characteristics as basically infantile, as HILZHEIMER (1928 and 1931) states regarding similar head conformation in breeds of dogs, cattle and pigs. HILZHEIMER's illustrations (1931) could just as well illustrate the status of the cat as that of the dog. In early adolescence, there is a clear tendency towards this head shape in all kittens to varying degrees in the different individuals. As they mature, these forms normally regress. The “grimace” of the Persian face is already indicated in some specimens of F. silvestris by virtue of the strong gradation from forehead to nose, and the characteristics acquired through domestication reinforce it. Psychologically it belongs to the pseudo-physiognomic, to which is genuinely physiognomic in animals. I have treated this contrast using the cat as an example (SCHWANGART, 1928), and here I would like to draw your attention to the noteworthy publications by Br. M. KLEIN (1930 and 31) with regard to the genuine physiognomics. — It must not be overlooked that HlLZHEIMER (1928) compared this tendency to “infantile appearance” with an equally domesticated converse, one in which the adult type exceeds the wild-type appearance. In this work, we encounter various evidence of contrasting domesticated types exceeding the standard norms of wild ancestors, and it is probably a common occurrence.
The “correct” Persian head introduces an excessively developed breed type. Nevertheless, normally exemplary specimens may still differ greatly, a sign of the significant plasticity of conformation in the cat. Compare fig. 4 with that of a French “champion,” fig. 5 in SCHWANGART and GRAU.
A peculiarity of the Persian breed is the different degree of expression of special domesticated characteristics in its head and face conformation depending on its colour. The most advanced step is the change of shape [literal: breaking the mould] – we can consider the average or “winner” of the test to be the blue (blue-gray), followed by the white, yellow and chinchilla, also “peach”, then the piebald; at the minus end are the black and the tiger- and classic tabby varieties. I think that is evident that breeding plays a part in influencing this order. In fact blue is such a favoured colour that some breeders think that “Persian” means Blue Longhair, and the Blue Shorhair is even spoken of as a “short-haired Persian” as a result. Likewise, traditional disregard for patterned animals, especially the gray ones, has meant their breeding has been neglected, doing them an injustice, as I shall show. However, I will briefly return to the discussion of the Blue Shorthair when discussing the blue colour, to show that in nature there is a tendency for colour to coincide with changes in conformation. Thereafter, the blue Persian should also be somewhat predictive in this regard. In any case, the practical goal of Persian breeding must be to bring all the colour nuances and combinations in conformation to the same high level as the blue Persian.
For my German Longhair breed (Fig. 5), in contrast to the Persian, this description applies:
“Slanting forehead, not pushed forward, flowing in a shallow arc to the bridge of the nose, or meeting it with only a very slight gradation. The bridge of the nose is elongated, straight or slightly hooked. The conformation is less stocky, the movement more fluid, and the tail a little longer than the Persian.”
In order to remedy the previously mentioned slander of the breed as “pointed-faced”, I have added to the description, the in both breeds, “muzzle is wide and not pointed.” By the same token, this same purpose is met by the remark: “The head complements that of the striped Shorthair.”
The German Longhair comes in the same colour and marking varieties as the Persian. For each of the two breeds, there are the following classes: Self-colour (black, white, blue, isabelline, and others), piebald (bicolour, three-colour) and “masked,” [tuxedo pattern] chinchilla, peach, “smoke cats,” silver, tabby (“tiger” and “marble” [classic]). I will explain in later sections the less easily understood terms in in this list. The German Longhair was officially introduced at the Berlin exhibition of the “Association for Cat Breeding and Cat Protection” in January 1930, after I described it in my “History of Breeds” 1929.
I decided to establish it alongside the purebred Persian for aesthetic reasons. For me, the conformation of the “perfect” Persian shows an excessive type with the strikingly deformed features already mentioned. It was also mentioned that this type, after a justified correction of earlier mis-breeding defects, should remain the only Longhair worth breeding. The recently established Birman breed, which is generally long-haired, cannot counterbalance the Persian, as it is also aberrant and is certainly not a pure longhair, as I shall show. But I often saw under our Longhair a form that bore near-natural features in head and face formation, resembling the wild, without violating the data and giving, in my opinion, a justified basic idea of a Longhair. I refer back to the differences between this, the German Longhair, and the Persian, and on the two common traits according to my outline presentation on “Breeding Classification and Evaluation Rules”. The head and face of the German Longhair bear traits of the northern F. silvestris without the infantilised domesticated features of Persians. With these eugenic considerations in favour of establishing the German Longhair come aesthetic satisfaction, and good specimens look pleasing by virtue of their balanced appearance. With my confession of preference, we must not cast aspersions on the so-called high-bred Persian, which is interesting and imposing with its excessive features. Infact the appreciation of his newly established German Longhair counterpart should contribute to the better breeding of those Persian colour varieties associated with poor conformation. Intermediate forms must disappear.
Convincing me ion this regard is the opinion of O. ANTONIUS (1922), that he made on two breeds of dogs that can be considered counterparts of the Persian and German Longhair in terms of their different head conformation. In the case of these dogs, the mistake is already lamented, but is likely to be repeated in the German Longhair if the lesson is disregarded by its breeders. ANTONIUS is considering (pg. 120) the great breeds of Swiss Mountain Dog as “ancestors of high-bred breeds,” namely, the St. Bernard and the Leonberger, and notes: “Unfortunately, this beautiful German Basset”(the Leonberger) “did not find the same following as its Swiss relatives, since . . . German fanciers were completely ignorant of its history. Instead of preserving as many as possible from once abundant stock, the dog fancy‘s verdict condemned them. Anyway, the Leonberger was the original. . . Its type is more faithfully preserved than that of the St. Bernard, which has recently been bred to be Mastiff-like, especially by the English.” The discovery of this passage in his excellent book was even more pleasing to me, because my knowledge of the Leonberger predates my advocacy of a German Longhair cat breed and in lectures justifying my project I had already made comparisons with the two dog breeds mentioned.
The thoroughbred breeding of this new breed is more demanding than that of the British or French-related Persians, whose pedigrees can bear comparison with those of the best pedigree dogs. (Unfortunately, the German breeding practice is often not even up to this task, often the expensively bought stock is bred down and then more is newly purchased). In the case of the German Longhair, it is important to create an elitge breed first. Only then can the breed gain a market value. The general shortcomings of our cat breeding already enumerated are pitfalls in the case of new breeds, something we know from other animal breeding disciplines. Breed clubs dislike recognising the products of a “competitor‘s” initiative. However, it is rarely possible to initially enter new ideas in an animal fancy in just one place.
With regard to different possibilities for the German Longhair’s conformation, I deliberately left leeway in my description in order to avoid excesses. Experienced breeders first decide to what extent and in which direction to commit themselves. Such points are: The hair quality, whether silky or more woolly, and details of the tail shape, especially if the so-called “Flag” or “Pleureuse” [veil] is required i.e. a very long silky hair curtain on a long, curved, sweeping tail, the hair maintaining its length along the full span of the tail. This form is also found in Persians as well as a stouter, woollier tail, which is probably better for that breed. Longhair questions are also addressed in detail in my “Notes on Cat Breeding” (1928/30) No. 7.
I have said that the “basic idea” of a Longhair, whether Persian or German Longhair, which I mentioned above in the common description for both, is not only traditional but is also “justified”. Among other things, it is also phylogenetically justifiable. In my booklet “History of Breeds” etc. (1929), and even in its predecessor (1928) I presented the view of a two-species ancestry of domestic cats. I connected this to R. I. POCOCK’s conjecture (1907a) that Felis silvestris SCHREB. (Fig. 7) is distributed in those parts of the Middle East and Central Asia from which the original “Angoras” came to Europe (allegedly into Italy in 1521, but not into France until around 1720). It is precisely this “basic idea” that corresponds in its main features to the conformation of Felis silvestris, even the inordinately slender and pointed-faced domestic cats more closely resemble variants of this wild-type (I’m thinking of those Transylvanian wildcats that can be seen at the time of writing in several zoos, including Frankfurt and Vienna), than they do to Felis ocreata G.M. (Fig. 8;), though this does not exclude the possibility of their crossing in the Asian regions where they come into contact with the Asian Felis ornata GRAY (Fig. 9) and its subtypes.
As previously mentioned, the standard Angora belongs to the moderate far northern wing of the Felis silvestris SCHREB. group, and the German Longhair remains within the bounds of the normal wild type, unlike the domestically moulded Persian. We know that “Felis silvestris” forms a great number of geographical types and, as I have already mentioned regarding the Siebenbürgen specimens, we can find more slender construction and narrower muzzles than desirable in a prize Persian or German Longhair; also we must add the Shorthair tiger-striped pattern (See Footnote). Such wildcats are then sometimes remarkably small, compared to their Northern counterparts; sometimes even smaller than some of our longhaired and shorthaired house cats.

[Footnote - This footnote will makes little sense to English readers without knowing that “rasse” means both “species” and “breed”.) Regarding the numerous subtypes of the very variable Felis silvestris SCHREB. and ocreata GM., very probably also Ornata GRAY, comes the view, as I have already expressed in 1928 (“On History and Typology of Breeds” etc.), that it is a member of the same Geo-circle. In addition, I have argued that F. silvestris and F. ocreata belong together in a common grouping and that there is also a fairly close natural connection to F. Ornata (I speak of F. ornata caudata GRAY).
In the meantime, B. RENSCH has published a significant work (1929), which especially addresses the problems involved and, incidentally, among its numerous examples overlooks the wild cats. RENSCH spoke of “geographic race groups” in which the actual “races” occupied their own ecological and seasonal conditions, and he expressed his concern over the use of the term “races” in animal breeding as it has a significantly different meaning that could cause confusion. First of all, stressing that this is not an objective difference, let me say that I agree with RENSCH in many respects. But as for terminology, I cannot agree with his concern over the different meanings of the same expression. I would like to see the term “race” restricted to its original sense of cultured races in its original sense. For the phenomena which RENSCH discusses, terms can be created with the help of the expression “type,” which also creates unity. There are then I.) Geo-groups containing Geotypes, II.) Species in the narrower sense contains eco-types with their own seasonal, physiological and other variants. In addition there would be (III.) the peculiar mix of mutations, “Offshoots of Races” (according to my mode of expressing “types”, depending on geo- or eco-types), that R. WOLTERECK has just studied (1981). It hardly needs saying at this point that the specific term “ecotype” does not come from me. As to the question of whether to exclude geographic race groups fundamentally from the species concept, as RENSCH wants, I am nevertheless in favour of extending of the species concept in a broad sense. Such species are then identified in the text as Geo-circles. RENSCH himself says (p. 12) that “in some cases a clear divorce between geographical and ecological race is not possible”, both are “conditioned by their environment”.]
There are significant differences when comparing the patterns of European long-haired breeds and the Northern wildcats. J. VOSSELER (1927) apparently found long-haired cats in Angora itself which are far from corresponding to the European breed standard. This particularly applied to their “smallness” and “delicacy” and more “elongated” body, peculiarities which I regard, like that author, as due to a “somewhat inadequate standard” there. These animals are nevertheless linked to F. Silvestris, like our good Longhair, having noticeably small ears and “low-slung build”. As a special attraction, the native race of these animals had fine silky hair - in contrast to our Persian - a “well-developed flag of a tail”, white fur - the author saw almost no other colours, a sure sign of deliberate selection, which prevented it from mating with the commonly found Shorthair - and disparity in eye colours - which would disqualify it against our standard.
Supporting the role of Felis silvestris as determining the conformation of pure long-haired breeds, the long-haired character must also be claimed. Out of all the available species, this one offered the best chance of successful selection for that trait, and certainly not F. ornata, the Egyptian form of F. Ocreata.
This claim of F. silvestris first of all contradicts the claim of the Manul (Otocolobus manul, and several subspecies, Fig. 10), which was considered a kind of strain of the Angora cats for quite some time. I have denied the possibility of this relationship (1928 b and 1929a) with reference to various strong differences between the Manul and the F. silvestris-ornata-ocreata group and this has been confirmed by a recent work by S.J OGNEV (1930) based on detailed material studies on the “Russian small cats” of the Manul group with the subspecies Otocolobus manul manul Pall. (1778) and O. manul ferrugineus OGNEV (1928) – together with O. m. Mongolicus SATUN. and O m. nigripedus HODGS. from Tibet - confirmed its genus and contrasted it with the other small cats inhabiting that area with the sole exception of the Eremaelurus thinobius OGNEV as a member of the genus Felis [Note: Eremaelurus thinobius was a synonym for Felis margarita – the Sand Cat]. There are two forms of F. silvestris, five of F. ornata GRAY, and the sub-genera of Felis, Prionailurus and Chaus, each with one species. Thus even the lynx-cat (Chaus) of the Silvestris-ornata-ocreata group is more closely related than the manul (see Footnote)).
[Footnote: OGNEV (1. c.) Regards the Manul, in comparison to the other wildcats mentioned here, as a “progressively” developed form of felid according to its peculiar cranial structure. The same tendency is also seen in Eremaelurus thinobius OGNEV, but to a much lesser extent, so that this cat remains closer to the rest while the manul is more distant form them.]
I do not personally have sufficient material knowledge of Chaus and Prionailurus. After the impression of several living and stuffed specimens, such as some of Chaus pelts (Fig. 11), it seems to me that this one should be ranked in its own genus. Also the only speciment of Prionailurus euptilura SATUN. that I have seen (living in Leipzig “Ipa”) greatly differed in physique and behaviour compared to F. silvestris, ornata and ocreata, which are closely grouped when compared to other small cats. On this occasion it seems appropriate to refer to my view, which has been explained in more detail in 1928 b, that F. chaus, contrary to the current opinion in cat books and magazines, has no claim to being a joint ancestor of our domestic cats. Incidentally, its claim is only asserted with reference to shorthaired cats. In a recent essay (1931 b), O. ANTONIUS, while considering his Schoenbrun F. chaus, explains that “it is quite incomprehensible how anyone could bring this form into the historical family tree of our domestic cat”. He states the importance of having knowledge of the living animal when evaluating such relationships. I particularly agree with this statement.
Referring to several such rejections of alleged ancestors and indigenous race formation, I have suggested (1929a) that “it should nevertheless be examined everywhere whether it is possible to distinguish indigenous types.” I emphasized this suggestion especially for East and Central Asia: “If literary or pictorial evidence of domestication of separate wild cats by Asian peoples is missing . . . this is not evidence against such domestication. There are plenty of wild species in Asia”. — If a supposition, which my colleague E. SCHWARZ verbally announced to me, is correct, even the Manul, whose relationship to the Angora is to be rejected, appears particularly unruly in captivity (see POCOCK, 1907 b) and among the Old World small cats is morphologically too distant from the root species of our Longhairs and Shorthairs to be ancestral to a special domestic cat occurring in its area of distribution. (Document in the Berlin Museum, collector C. WACHE). I have not yet seen this piece of evidence [See Footnote]).
[Footnote: The “unruliness of the Manul” was noted by POCOCK when comparing it to examples of both F. silvestris and F. ocreata. The frequent general judgments about the degree of taming of small-cat species are often obtained from a few, or even single, specimens and then do not take into account either age differences or individual temperament, nor the crucial factor of previous treatment of the animals. The frequent general judgments about the degree of taming of small-cat species are often obtained from a few, even individual specimens and then count neither age differences nor the individual temperament, nor the crucial factor of pre-treatment of an animal. It may be assumed that the same practical knowledge of different types of the Silvestris-ornata-ocreata group can be gained, as POCOCK already speaks of the trusting nature of the specimesn of F. silvestris that he keeps (i.e. certain individual ones) and has already mentioned to NEHRING (1888) the case of a perfectly tamed wildcat raised by ALTUM. Of course, there are certainly differences in taming between small cats. While it is fairly sure among the Ocelot group, Serval and Lynx, we see the Caracal, which can be used for hunting, tethered in the courtyard by its owners in India because of its univeral fierceness, and in zookeeping experience, the unruliness of F. viverrina seem unanimous. POCOCK's statement about the Manul should not be universally accepted as this cat is rarely seen in zoos. As an example of the numerous contradictions in judgment on the behaviour of small cat species in captivity, here is a comparison between the statement one of A. BREHM’s sources and the personal experiences of the well-known explorer E. V. PFIZENMAIER (1929) on the relationship between tamed large lynxes and the domestic cat. BREHM notes an indomitable persecution fury, but also a pattern of house-tameness, among the northern lynxes. PFIZENMAIER makes this judgment at least for Lynx pardalis orientalis SAT. in the true sense, as specimens of both species, when brought together young, formed a lasting friendship, and it was particularly appealing to observe the gentle manner in which the lynx played with the tomcat. There is a picture of the pair playing. Therefore it is conceivable that members of larger and smaller cat species would have to agree with each other just as we are used to in big and small dogs. Given the domesticability of larger species of small-cat, this view is significant.
I have already shared my view of the close relationship of the “species” F. silvestris SCHREB. and F. ocreata GM. into a geological circle with a considerable number of two-way geotypes, distributed over a vast space across three continents of the Old World together with part of its archipelago, (1.c. 1928, 1929). Extremely developed forms are undoubtedly found in the climatically most remote zones, for F. silvestris northward already in our region, for F. ocreata etc. in East, Southwest and South Africa. I was especially struck by the fur character (extreme shaggy or woolly and “shorn”, velvety), the overall size, the tail length, the height of the legs, the ear size, features which can be discerned from skins and from good taxidermy specimens and especially in living animals, where details of conformation of the head, face and other parts may also be seen. Likewise, I sought to assert that an exclusive distinguishing feature of species status does not exist.
There are two contact zones. One is a direct contact zone in Asia and one is across the islands and peninsulas of the Mediterranean and is broken by the sea. The pelts that I examined at the Berlin Museum was scanty for the continental contact area. At least, it showed no fluid transitions. Also, the museum material highlights how much the tropics are favoured by collectors, compared to often crucially important specimens from closer to home.
More revealing is the distance of indirect contact between the northern (silvestris) and the southern (ocreata) group of the species-circle. In earlier work (1.c. 1928, 1929), I tried to explain how the formation of special insular types could not have masked the general impression of an intermediate character.
Furthermore, even among the rich African material available in the Berlin Collection, there were noticeable differences in fur character and tail length. Thus, of the specimens from Togo, Angola, Southwest, most had fur that was not velvety or “shorn,” but had more straw-like hair, but there was still a considerable difference between them and our silvestris material. It can hardly be attributed to a mere seasonal difference. Most pronounced is the deviation from the ideal ocreata type in Northwest African specimens, where the generally longer hair on the tail become slightly bushy, and the tail length probably exceeded that of the northern F. silvestris, but was less than the average length of our Shorthaired domestic cat. In the case of head width and face length between the wildcats, there were such great differences that one must reckon with regional difference or significant ecological differences, as I have already seen previously in the few zoo specimens. For studying African wildcat material, I recommend a collective method that takes the differences in elevation into account. You may then discover wildcats with a slight tendency to the F. silvestris physique and behaviour at higher elevations in central and southern Africa.
Since my publication in 1929, a communication from ERNST SCHWARZ (1930) on “The Wildcats of the Balearic Islands” has made the important addition of F. catus jordansi E. SCHWARZ to the European-Mediterranean wildcats. It is closely related to the North African “F. catus mauritana” CABRERA. An overview of the Mediterranean types brings the author to the same opinion of the “species” that I first expressed 1928 b; I prefer to consider them a Geo-circle corresponding to a meeting of F. silvestris and F. ocreata. “It does not seem feasible today,” says SCHWARZ, “to speciafically separate the African wildcats of the. , , ocreata group from the larger, longer-haired sivestris group.” [See Footnote]. In connection with E. SCHWARZ’s conclusions, I refer to my communication regarding the “Riviera” cat in my 1929 brochure, in the section on Mediterranean forms. It is a stuffed specimen in the Nice Museum, closer in shape and fur type to African wildcats than to European ones. To exhaustively examine it, it would have to be dismantled. This specimen joins the Balearic cat and introduces an ocreata-type onto the European continent. A wildcat is still sporadically present in the Maritime Alps, whereas the lynx known there is said to have been eradicated.
[Footnote: The name of the new local form would have to be in accordance with the naming method I have followed F. ocreata jordansi. According to POCOCK'S findings (1907a), LINKE described a short-haired classic tabby domestic cat as F. catus, believing he had a wildcat in front of him. Catus can not be considered the species name of some wildcats instead of ocreata. I mention here this nomenclatural issue, otherwise the significant fact that the new local form belongs to the African group, which I summarize as “ocreata”, while a number of other Mediterranean island types are closer to the “silvestris” group, would not be clear. SCHWARZ also offers a geological interpretation of the contrast between this wildcat and those of other Mediterranean islands.]
Thus, first of all the groups F. silvestris and F. ocreata are combined into one single circle of geographical types (a “geo circle” with “geotypes”, with some interposed insular specialities), thus an overview of the whole, in its hitherto accessible horizontal variability, gives a clear confirmation of the “BERGMANN Rule” as well as the “ALLEN rule” (to apply the name suggested by RENSCH (1929) for the second proposed name). RENSCH has given both of these a detailed analysis, and in particular has analyzed the exceptions to the former and extended a rule originally for warm-blooded animals, to the field of cold-blood animals. BERGMANN's rule states that “of closely related warm-blooded creatures the larger live in colder areas, the smaller live in warmer areas.” This obviously applies to the whole of our circle, as well as to the F. silvestris grouping. The African wildcats are still poorly researched. According to ALLEN's rule, “in species in a racial circle [species family] living in colder climates” (he uses RENSCH‘s terms), there is a relative reduction of the extremities, a reduction in ear size, and shortening of the tail. The applicability of this rule to our Geo-circle proves a comparison between its northerners and Africans. The reduction of ear-size, a slight shortening of the legs and a considerable shortening of the tail is most pronounced in silvestris, along with lengthening of the hair. It is most probable that climate has played its part in the same direction incrementally within the species’ range. Investigation into this, taking elevation into account, is desirable. The African cats were already characterized as “large-eared” when compared to European wildcats.
Regarding the development of these differences, I agree with RENSCH's assumption that it is a functional adaptation that has become hereditary. With regard to causation, he reprises this idea, which I also understand: “The internal temperature of exposed body parts (especially the ears and tail) is in all cases considerably less than that of the trunk, and this depends to a large extent on the outside temperature. A growing ear or tail will therefore become different in size as a result of the known effect of temperature on physiological processes, depending on the external temperature.” This may perhaps also be based on the fact, mentioned by the same author, that warm-blooded animals in their youth undergo a “poikilothermic” period during which “a substantial part of their growth takes place or the development base for further growth is laid down.” — RENSCH adds that clarification of the principle of this “somatic inheritance induction” [see Footnote] is needed “to solve the problem of how such differences can become hereditary”. In a different passage of his book, he then correctly counters the widespread tendency, in the case of the lack of an answer to “somatic induction,” to justify the “principled rejection” of the case, that “the part of the physiological Chemistry, which deals with the chemical relations between germ and soma, must be overwritten with an 'ignoramus'. This ignoramus applies to events that have been experimentally proven, such as those that can be derived from circumstantial evidence, and not merely for phenotypic induction.
[Footnote: I drew attention to the character of the term “somatic” induction as pars pro toto [“one part repesenting the whole”] (SCHWANGART and GRAU 1931), since psychological things are also inherited, and applied the term “phenotypic” induction to this principle. The first term, “somatic induction” is also suitable for the sub-area discussed here.]
Unless the signs are deceptive, we are close to experimental evidence of phenotypic induction, however major natural difficulties stand in the way. At first, however, it seems important to emphasise that it is inadmissible to deny the validity of signs of an effect, in this case of a causal connection to phenotypic induction, just because the question is not cleared up for the documented events. In this respect, the same rule applies to circumstantial evidence as to experiments. In the sphere of daily life, no matter how carefully we judge the accumulated circumstantial evidence that exists in support of somatic induction, we have sufficient belief to arrive not only at acquittal, but possibly a conviction. I find highly questionable that breeders dismiss phenotypic induction as "scientific absurdity" equivalent to superstitions such as the "maternal impression." To me it seems that the common good of breeding practice and breeding biology requires a planned continuation in the such as RENSCH (1 c.) started to determine the possibility and degree of probability of phenotypic induction by classes of instances (such somatic and some psychological in nature). And in the case of pet science we must use the practice for such investigations and similar trials. From this field, beautiful examples of circumstantial evidence of inheritable and hereditary somatogenic alterations were presented by H. Baum in his Rectorial Speech (1931).
You should never make a pronouncement based on "circumstantial evidence." A court is tempted to do this, for instance when it passes death sentences on the basis of circumstantial evidence. The science itself contained and still contains numerous cases which presupposes a need for evidence.
With regard to tail length in wild cats, a parallel investigation into the lynx is recommended, on the one hand those in the Arctic and the North and South temperate zones, and on the other hand the tropical caracal with its significantly longer tail, and a comparison between these in particular. According to OGNEV (1. C) the special position of F. chaus as "Catolynx" (which, as well as tail length, was also based on the dentition being intermediate between Felis and Lynx) is shaky. I do not know to what extent a closer comparison has taken place into tail lengths among specimens of this relatively uniform, relatively short-tailed cat occupying widely different climates in a distribution area extending from East Africa through Southwest and Central Asia to India and Southeastern Europe.
Continuous transitions between "geographical races" (geotypes) are considered by RENSCH (1929) "as a criterion of fertile crossing", which is probably self-evident. On the other hand, fertility does not have to exist between such extremely endemic types. I put forward (1928, 1929) arguments against the traditional view of the infertility of the hybrids originating from crossing F. silvestris and ocreata (respectively with domestic cats). Since so much speaks for both being a unified species, it should hardly require such evidence. However, the test was a worthwhile experiment, and even the far-flung and form-dissimilar types of this geo-circle, such as the Scottish F. s. grampia MILL. and the fine-haired types, particularly the high-legged South African wildcats whose pelts are in the Berlin Museum (which I reported in 1928) also produce fertile hybrids.
In our domestic cat stock there are moderate and extreme forms in both directions, both towards ocreata and silvestris types. The second (silvestris) direction includes the two long-hair breeds already described - see their common descriptions earlier. Certain Shorthair varieties resemble the first (ocreata) direction, which will be mentioned here, while another of the Shorthairs, which will also be discussed, approaches the silvestris type in general type and in its details and may even be the same as it. Considering the claim for silvestris as the basis of the Persian and the German Longhair, it is noteworthy and also supports the placing of both clans in a single type-circle where they have developed the same domestication pattern and colour qualities in both long and short hair types. According to previous experience, there is unlimited fertility between all the form-types of Longhair and Shorthair.
Here I refer to one of the three groups of wild cats closely related to each other and to the domestic cat, F. ornata GRAY (Fig. 9). I start from the boundary given by OGNEV (1930) by annexing F. caudata GRAY and separating (with LÖNNBERG) F. pallida, which was assigned by A. BIRULA to this group, into the separate sub-genus Podiaelurus. According to OGNEV, the distribution range of this considerable series of subtypes - 5 South Russian – has reached a considerable part of southern Russia, Mesopotamia, the Kerman province of Persia, Afghanistan, and reaches deep into India. It is a "steppe cat" and probably deviates from the "forest cat" F. silvestris in their two shared areas. On the other hand, she will encounter the Manul. According to OGNEV, it goes vertically in southern Russia from about 600 to 1000 feet (whereas silvestris exceeds 6000 feet). My idea of the animal is mainly based on the photograph from the HAGENBECK Company reproduced here. Thereafter, F. ornata is very similar to the silvestris and ocreata groups. While the hair comes closer to the first type - OGNEV finds "the winter coat at least being dense and bushy", and even the summer coat cannot be really short-haired according to his indications, and F. ornata caudata also comes with a "bushy tail" - and even the relatively "short tail" points to F. Silvestris, while head and face formation seem more akin to ocreata.
The possibility of breeding a long-haired cat from F. ornata seems to me to be given. (See the somewhat long-haired variant Fig. 9). One might imagine that the relative slenderness and daintiness of the Angora, described by VOSSELER (1927) in the city of the same name, was connected with such an origin. But assume there would be a further import, which is credited by its name as "Persian". The tendency of our Longhair to strong growth, stockiness and other silvestris-like features, however, suggests selection for delicacy on the silvestris material,a trend that is seen in all luxury breeding, as well as in ours, which now wishes for a "dwarf Persian." Furthermore, F. silvestris lends itself to the selection of pronounced long hair alone. - The participation of F. ornata is not excluded. In places where both are found, it would hardly have made any difference in the making of domestic cats.
In order to differentiate between F. ornata and silvestris, skull features are shared (OGNEV), as is the case between typical silvestris and ocreata, and even for the differentiation of subtypes of F. ornata minor features of this kind are used. OGNEV records an indication by N. A. SMIRNOW of a "hybrid" between F. ornata caudata GRAY and F. silvestris as being possible. Likewise, transitional forms between the two are conceivable. If, according to the present material between F. silvestris and F. ocreata, there is a pronounced geotypical relationship, then at least silvestris and ornata are also ecotypically related to each other. Fertile hybrids will be achievable between ocreata and ornata, the same as between silvestris and ornata.
The English breed book "Domestic and Fancy cats" claims (p. 46) cases of fertile hybrids in the field between "F. ornata" and the domestic cat, but gives no evidence and assures us of a similar hybrid with the "spotted wild cats", for example F. bengalensis, which is probably too far away from our Form-circle.
The wildcats I listed in 1928 and 1929 as "Felis caudata GRAY" are now known as follows: F. ornata was morved to the higher-level ornata caudata category, F. pallida BÜCHNER was moved into its own subgenus Podiaelurus (see above), F. chutuchta, not mentioned by OGNEV, but according to BIRULA's original description (1916) it would be close to pallida, since the author sets it apart from them; homeland is South Gobi, material is a pelt specimen; F euptilura moved from the true ornata into a separate subgenus prionailurus; F microtus MILNE-EDWARDS is F (prionailurus) euptilura microtus M.-EDW., F. mandchurica MORI is close to OGNEV F. euptilura. Afterwards ornata and thus our cats, which belong to our great geo-circle, seem to spread east to the Pacific Ocean. Towards the same region, species differ from them, one of which (in the subgenus Podiaelarus) is still very close to the silvestris-ornata-ocreata circle, while the other (Prionailurus) is considerably further away from it.
In a discussion of our long hair you do not get past the so-called half-angoras. R. C. BAMBER (1927) summarizes the rather poor results of previous hereditary scientific experiments on the crossing of long and short hair: “ From all useful data it can be seen that the 'Persian' or 'Angora' trait is a simple Mendelian recessive. But it must be admitted that our knowledge is imperfect. In particular, nothing is yet known about the inheritance of the gradations between fully long hair and fully short hair". From the point of view of our purebred breeding, we must first of all note that all these experiments dealt only with the Angora and its shorthaired counterpart, without taking into account further characteristics of long and short hair, which are just as important for us. The examples which BAMBER raises of "gradations" between distinct Longhairs and Shorthairs all stem from frequent reports of supposedly local variants, of which those that were open to examination have proven erroneous. The same variants are also present in our mixed breeds. We have the shape of typical and atypical long-haired cats with different length fur, also different type of fur e.g. woolly or silky hair, and among those which, according to the overall impression are considered shorthaired, we find velvet, rough and straw hair. All of these, as we previously mentioned, have precursors in the ancestral wild types and are transitionally related to them. Mrs. BAMBER rightly recommends the naked cats for use as another partner in heredity experiments. I have reported on such cats in SCHWANGART and GRAU 1931. Mrs BAMBER is also mistaken in view of this phenomenon being localized ("Mexico").
In hybrid litters of thoroughbred Longhairs and both velvety and coarser Shorthair (I did not happen to see straw-hairs involved), there was indeed a division in hair quality. In addition, in the only such litters – which I disliked of course - where I saw the material worthy of further evaluation, there were no intermediate forms, in accordance with TJEBBES‘ scheme (1924) as reproduced by Bamber, so that here too the Mendelian scheme of inheritance was present.
Nevertheless, there are cats that appear to be real "half-angoras" [Semi-longhairs], cats which have intermediate hair length (which one otherwise does not find on domestic cats, the norm being a distinct gap between straw hair and long-hair) and at the same time having other formal characters that are intermediate between Longhairs and Shorthairs. Of course, it should not be said, or considered probable, that they are "intermediate" in regard to all of the other characteristics. On the contrary, I have seen quite different combinations.
From the point of view of pure-breeding, we first conclude that the "half-angora" is generally excluded from gaining prizes at shows (and this is far from being the case everywhere), and we warn against haphazard crossbreeding attempts between Longhairs and Shorthairs. After the last experiences, however, two special cases require special attention.
After I returned from a trip to the French Riviera in 1928, I gave a report at a meeting of the "Association of Cat Breeders" (Dresden) about a certain type of "half-angora", which I frequently noticed there (in and around Cannes, in Juan les Pins, in Nice). I saw the same type this year (1931) in Paris where I was judging at the International Cat Show, not at the show, although "half-angoras" were allowed, but in the inns and shops. WI also found this charming type in Northern, Central and Southern Italy (Naples, Ischia), but not in our own country. I described it in my French exhibition report:
"They are low-set, rather elongated small animals with elegant movement, and a short face of a distinctively special shape that looks exceedingly cute. Its hair is silky and slightly less than half-long, except for the tail, where it forms a veil. In essence, all the specimens I encountered were gentle, vivacious and intelligent. I recommended to the management of the "Societe centrale feline" to check whether this type is consistently inherited, as it appears. If so, this may be considered a breed."
Obtaining a non-segregating [pure-breeding] "half-angora" original type of long- and short-hair would probably be pleasing for the sake of its psychological advantages, for what I noticed regarding this strain can be said of Longhair-Shorthair mixes in general. They are said to combine the virtues of both parents, the quiet nature of most long-haired cats, without its frequently seen apathy, with the livelier temperament and quick mind of the Shorthair, without its inherent restlessness.
The second notable special case of "Half-angora" [Semi-longhair] is that of the new "Birman" breed (Fig. 6). At least I consider these beautiful animals to be a product of crossbreeding, namely of the Persian (of suitable colour) with the Siamese. To me, this interpretation seems obvious regardless of whether the ancestral animals really came from Burma or were produced in France itself. (In order to recommend the breed, it is surrounded by similar legends as the Siamese - sanctification and breeding in temples, acquisition by robbery and risk to life, etc.).
The original description ("standard") of the characteristics of the "Birman" is ("Revue feline de France" [Feline Review of France] 1930):
"The body is elongated, well-proportioned, set low. The head is short, with large ears, the forehead prominent, the nose curiously tilted upwards. The eyes are large, dark blue, iridescent, with a gentle expression. The hair is quite long, parted on stooping. The tail is very long, thickly bushy, forming a broad flag giving the animal the appearance of a squirrels." (This comparison is an exaggeration.) "The colouring is light cream with golden tones on the neck. The mask, tail, ears and paws bear the same deep brown as the Siamese, but the brown, 'booted' paws have pure white toes.
They are difficult to rear, perhaps as a result of inbreeding, because until now in France and perhaps in Europe only a single strain of this breed is known.
These cats are very affable, clever, genial and affectionate. They jealously cling to their master and can not bear to be separated from him.
Some connoisseurs see in the Birman cat the ancestors of the Siamese who sprang from a cross between it and the Annamese cat."
The aforementioned problem of rearing Birmans are shared with the Siamese, and these are surrounded by a superstitious-misunderstanding type of treatment, - mollycoddling by keeping them warm, depriving them of fresh air because they are considered "tropical animals", favouring serious hereditary defects (see SCHWANGART and GRAU), - as well as a form of albinism that leans towards flavism ("Siam dilution"), which will be discussed later. The Siamese may also have passed inbreeding defects to its Birman descendant. (I do not know whether the Birmans share the Siamese peculiarity of being born white and gradually developing its colour.)
The assumption of Siamese originating from the Birman is, as already indicated, the wrong way round. A look at Fig. 12 and Fig. 4 shows the Birman is actually a combination of Siamese and Persian. To begin with, the Birman has the same colouration, except for the white toes, which are probably due to a whitening factor from the Persian side and are not exactly welcome. The semi-long hair corresponds to the "half-angora" trait, and the tail formation corresponds to that of the Longhair. The head and face form have pronounced Siamese and Persian traits, the overall shape is similar to that of the French half-angora variety, which I described earlier. The eye resembles that which is peculiar to the Siamese.
As for the "Annamese cat" which, according to the interpretation of the Birman given above, produced the Siamese, from the vague information one suspects that it is itself a Siamese descendent, probably a hybrid with a Blue Shorthair or Blue Longhair.
For a detailed description of the Siamese (Fig. 12) I refer to my "breed categories" (SCHWANGART 1929b). With this old recognized breed we now reach the Shorthair breeds. It is one of the exceptional cases in which conformation criteria were used before the appearance of my racial system in Shorthairs.
I reluctantly decided to recognize the Birman as a breed. Such reservation should be appropriate. Reliable authorities (the directors of the "Societe centrale feline") assure me that its captivating appearance is consistently inherited. In addition to the pattern, which is shown in our picture, the exhibition also included "impure" Birmans (sang impur) and so-called "Thibetans", which were little better than those. These animals betrayed the Persian blend in varying, even less appealing, combinations. Local Shorthairs may have been involved. It will be important to maintain it to a breed standard, and this also probably depends on whether it is a new breed or, if it really originates from Burma, if there are successful imports of additional bloodlines.
With the Birman and also with the French "half-angoras" described above, if they are genotypically established, we find breeding products that originate from crossing established breeds. This procedure, which is followed in all kinds of animal breeding, is generally not recommended in cat breeding. At the present stage, it is important to produce, from the mixed stock, distinctive types worthy of being breeds; currently have only a few of these. A preponderance of the admittedly more convenient attempts at crossing these few existing types would amount to a leveling which counteracts the necessary pursuit of forming distinct breeds. Even in dog breeding, which has progressed so much further in creating good, sharply differentiated breeds, the recent addiction to crossbreeds between established breeds is beginning to show serious consequences.
In the Longhair breed we saw the body form as the key characteristic deliberately selected, initially in the design of the standard English Persian. By putting German Longhair to one side, I proceeded in the same way. With the Shorthair, a characteristic conformation has played a part in addition to colouring and the arrangement of colour in the Siamese. We recognise the same combination in the semi-longhaired Birman, which one could call Longhair Siamese if not for modification of form and features caused by the Persian. Except for the Siamese, the Shorthair (the collective name for so-called "European" or "ordinary" Shorthairs, "domestics" as opposed to so-called "purebred" or "high-bred" cats) that occurs in the English breeding trend, which is followed the other nations, can trace back its conformation.
Nevertheless, there are two notable cases where a consideration of conformation occurred: the so-called "Abyssinians", also known as "Nubians", and the Blue Shorthair. I will discuss these cases immediately. Apart from these, the attempts to create precious breeds out of the richly differentiated stocks of the "ordinary" Shorthair in due appreciation of its high beauty shows a one-sided appreciation of the play of colours; this is reflected by the founding of more and more specialist clubs for individual colour variants. Shorthair breeding is now emulating Persian breeding with clubs for “Black and White”, Blue, Chinchilla, “Silver and Smoke”, Orange, Cream, “Tortoiseshells”, “Brown tabbies” etc. The difference is that in the Persian all these colour variants are subordinate to a particular breed conformation, whereas in the domestic short hair they are supposed to be a substitute for breeds, and therefore the "middle ground" is considered the ideal form and is the goal in order to achieve high honours. In reality, however, the show specimens - note the pictures in English breeding guides! - only rarely approach that intermediate ideal. In fact the impression of the "middle ground" most often stems from one feature tending in one direction and another tending to the other extreme. Quite often the features of show specimens are surprisingly disharmonious combinations. If, from this viewpoint, we examine the appearance ofthe wildcats, it is difficult to find cases where they are so blatantly disharmonious. This effect means, depending on how we measure it, we can claim to shape natural objects, as well as those made closer to home. Even here the impression of harmony varies. A combination of flat broad forehead with a strongly tapered muzzle, as in F. ocreata, or of a slightly domed, broad head with a slight tendency to a pointed face, as in southern F. silvestris and (Fig. 9) in F. Ornata can, from a human perspective, hardly be considered strictly harmonious in form.
The ever new fashionable colour specialities for Shorthairs line up like an "inexhaustible" play of colours in the existing spectrum of cat breeds. In addition to preferential groupings (which we also do not want to set aside) such as "masks" (breed-based in the Siamese!), "Tortoiseshell" (black and red-yellow shades without white), "Spanish" (as before, but with the addition of White). In the "tortoiseshells," the sex-linked inheritance of their colour combination is considered a "breed feature" in an otherwise aesthetic rating, and is not allowed.
The asset of the English breeding methodology is their never-ending concern for the quality of hair and colour, i.e. the distinctive character of the hair, the sheen and the depth of colours. For our part, we must not forget the importance of colour and beauty in cats in our pursuit of useful race characteristics, and we must also cultivate those aspects.
I mentioned two cases where the English approach was not quite able to ignore the particularities of the respective local Shorthairs to disable those major factors. One is the case of Blue Shorthair. The so-called "Maltese dilution" ("blue", ie blue-gray, which is "diluted black") has, as I previously mentioned regarding the blue Persian, long held breeders spellbound. Blue shorthaired cats appear frequently in some places, such as in the Black Forest, in Switzerland, in parts of France, and depending on whether cats were kept as pets or on a more industrial basis, we would presume them to be bred either by cat lovers or by fur-breeding concerns.
The English cat fancy now found it necessary to grant the Blue, alone of the domestic Shorthairs, two form variants, which they called "British Blues" and "Russian Blues" (colour nuances will be listed later). (See, for example, Figures 24 and 54 in Morton's "Domestic and Fancy Cats"). The "British" is heavier, more round-headed, but overall is still the English "middle form" associated with Shorthairs. The "Russian" (whose name, like most claimed countries of origin, is a misnomer – one sees these "Russians" particularly often in Mediterranean reagion) are slimmer and more pointed-faced. How to classify all the intermediate forms that form the majority remains unclear. The valuation principles vary, for example the breed book by FR. SIMPSON (1928) recognizes only the slim variant. It seems certain to me from my own experience of the Blue Shorthair that a relatively high percentage lean towards the slender form. It also has many animals that correspond to my well-founded "slim breed". (Description of this breed appears later; also see Fig. 23 -24). Only black, which is not so cherished by English breeders, is as numeroius as the blue. Where the blue ones correspond to my "slim breed", I classify them as such. In England, they appear among the "Russians," along with cats that do not lean so far towards the slender form, and with specimens that approach that category in some feature or another.
The exceptional position of the English separately recognising the slimmer type of blue shorthaired cat (the "Russians") is based on two causes; one is the particular attention paid to that colour compared to differences in conformation, and the othe is the tendency of blue to cause a slimmer body, head and face. The existence of a stocky build of Blue Shorthair has already been explained by a deviation from the form in the older Shorthair race, F. ocreata GM., In addition, in striking cases, it can be explained by the co-paternity of F. silvestris. In this case, the Blue shorthair does not differ from other colours of domestic Shorthaired cats, and we find no reason to assume this has any special tendency towards excessive conformation in the opposite direction from the slender form, which is also differently coloured. As a whole, it follows that blue hair tends to deviate in intensity in opposite directions.

Consider Fig. 13 and Fig. 14 showing two unrelated representatives of this colour. The contrast to the slim variety is surprisingly pronounced here, with the head shape approaching that of a mediocre Persian. A crossbreeding with the Blue Longhair, which cannot be strictly ruled out, is unlikely in these two cases. For the cat in Fig. 13, a number of ancestors of both parents’ lines are known, and including a Black Shorthair, while the rest are all Blue Shorthairs. The kitten in Fig. 14, a foundling, only the Blue Shorthair parents can be identified. In its head shape, it is above the lower limit of the infantile-Persian type which I have mentioned as a rule in cats in early adolescence. The picture shows the animal when about four months old when the juvenile form should already be reduced. The third example is that of Fig. 4 in SCHWANGART and GRAU (1931), where development has tends towards the pug-face. After all, in the case of the Blue Shorthair among the heavy, round-headed and broad-muzzled cats, it is exceptionally possible to find the characteristics of the Persian head, exceeding the form desired for the German Longhair or the shorthaired tiger tabby in Fig. 5 and Figs. 20 and 21. The fact that the more excessive forms are noticed more often, is evidenced by a term used by German breeders: "Shorthair Persian".
(Such a name is outlandish. Apart the Persian name already being assigned to a long established purebred type of Longhair, there is the danger of giving an incentive to create a "short-haired Persian" through the haphazard crossing of Persians and Blue Shorthairs is out of the question).
Some cat books at home and abroad combine the appearance of North American blue cats with a supposedly special hair quality, slightly longer than our Sshorthair, and sometimes "short-haired with frill", so that one thinks of half-angoras. We find these called "Carthusian" or "Maltese" (after "Maltese dilution"). The splendid collection of Blue Shorthairs at the Paris cat show this year (1931) were cautiously titled "said to be Chartreux" in the catalogue. They did not include any American specimens. What could be seen were a variety of shapes of pure Blue Shorthairs. It is, therefore, quite possible that the name Carthusian is in reality not assigned. It would be useful to breeders who want to select for the Persian type in Blue Shorthairs (like Fig. 13 and 14) (see Footnote).
[Footnote: Among the few cases of completely Persian-type heads on Blue Shorthairs which I have seen, there was recently a verifiable crossing with a Longhair. Caution should be exercised when assessing such animals.]
Why the Blue Shorthair has the same tendency to contradictory shape development does not apply to the blue Persian is explainable. If the breeding direction which was extensively devoted to this colour was severely opposed to slender features in the Persian, then so also were the breed foundation cats since Felis silvestris naturally tends to stouter forms. Thereafter, this corresponds well to the case of the Blue Shorthair, even in the Persian the blue seems to favour excessive form (as I showed in the section about this breed), but in that case in only one of the two opposing extremes.
This brings us to the idea of combining modifiability in the forms of blue, which has a tendency to extreme types, with the character of blue as a "diluted" colour. Among the preferred domestic colours, the "diluted colours" are also extremes. I will return to “dilution” and its tendency to upset the structure, to weaken the construction.
The second kind of native Shorthair, to which points are awarded for conformation in England, are, as mentioned, the so-called "Abyssinians" (often called "Nubians" in our country). This purebred breed was cultivated by the recently deceased English breeder H. C. BROOKE. His monographic study (1929a) shows that his model animals (Figs. 15a and 15b) were selectively bred from English stock for decades, and not from imported stock or descendants of imports, as is claimed and as I had originally believed. In colour and fur quality, however, these specimens resemble the furs of those groups of East and South African F. ocreata which I examined in the Berlin Museum Collection (see above, also SCHWANGART 1928b, 1929 a). BROOKE himself chose the name "Abyssinian" for that reason. His instructive illustrations right at the beginning of his study, depicting a strongly broken-striped, slightly straw-haired and relatively stocky built "Felis caffra," next to which is a finely mottled cat which is only striped on its forehead, tail and part of the legs, a slimmer, velvet-furred "Abyssinian Type" of ocreata (his example of a perfect specimen). This proves his knowledge of the geographical variability of African cats.
According to the breeder's wishes, the breeding line should also correspond in shape to the depicted wild representative; the original standard requires:
"The breed is very delicate and very elegantly built, with delicate slender legs, an elegant head, quite large ears and sparkling eyes. The so-called 'British type' [refer to Blue Shorthair, above] of breeding stock is out of place here. We also do not want a short and round head, nor small ears, and still less do we want a heavy build and bulky legs. "
In the main, then, the ideal "Abyssinian" corresponds in shape to the previously described "Russian-Blue" English standard. However, this ideal was not achieved by BROOKE’s specimen. In this they are both more like the wildcat reproduced here in Fig. 8, which represents a somewhat lean "hybrid". So you are already getting closer to the "British type". In comparison to my "slim race" (see Fig. 23, 24), which we often compare them with, these role models are distant. By the appearance of the two BROOKE patterns pictured here in Vienna, my belief in "Nubian import", which was then claimed (but not by the breeder!), was disturbed by the relatively short, strong tail, especially by “Bun.” (In the picture “Ras Tafari” appears more stocky than he really is). Otherwise, as I said, the image obtained by breeding is perfect.
Selecting for this type would be just as possible from continental stocks. In the shows and everywhere else, one encounters among cats - albeit rarely - animals that come close to this in every way; those that correspond to it in individual details are not uncommon. So it depends mainly on the hair quality and colour. Our comparison among animals, which correspond to the requirements, further expands the scope for the figure.
There are certain interesting colour and hair type degradation phenomena. These became known to me from BROOKE’s breed itself. Under a Viennese collection kept in a cattery and in another in Dresden, which was also carefully kept, a sudden change to “red” appeared. At the same time strong stripes appeared on the Viennese individuals, resulting in a "red tiger" pattern, while among Dresden cats there was a specimen that was almost solid, rich "red" with just a little hint of stripes as in the wild-coloured mottled ancestors (a rare nuance). So here the change only concerned the colour and not the markings. In a littermate of this red cat, parts of the coat were slate-gray, and the striped pattern had increased in size. The mother of these individuals had a pale yellow tone that outweighed the characteristic mottling, and the hair character, as a result of its lengthening, almost entirely lost its velvetiness.
In the treatise SCHWANGART and GRAU (1931) I assigned the "Abyssinian" to the "degenerate forms" of domestic animals, following the concept of ÄDAMETZ (1926) according to ÄRNDT, as an example of a "reversion to the original form", according to the colouring and hair quality, and as a "throwback" (pg. 247). There are several possible interpretations: genuine atavism ("spontaneous atavism"), which can affect both individual features and the whole appearance, crossbreeding atavism, and finally a simple survival of some of the domestic cats in this particular wild state would be an option especially since this interpretation certainly applies to the tiger pattern and its variants as well as to the nuances of wild colour. However, considering the centuries-long separation of our domestic cats from their African ancestral land, I consider it unlikely that they would be able to perpetuate an “imitation" of the Nubian F. ocreata coat.
If, in the case of the "Abyssinians," crossing atavism applies, the (exceptional) occurrence of this reappearance of wild colouration in domestic cats would also be reasonable among its remaining variants whose existence is usually based simply on survival. With regard to the structure of the wild-type coat, it is argued that the "red" (which are nuances of deep orange to cream-coloured patterned and self-colour) cats are derived from streaky gray due to loss of the black pigment (see, for example, VAN BEMMELEN 1931). The rules of genotypic behaviour of wild colour and wild pattern are given in my sections on pattern and colour.
Particularly with the change of the Nubian colouration into "red" and more pronounced striping one reaches a comparison with old evidence of domestic pet colouring developing in the cat. The well-known ancient Egyptian picture "Cat eating a fish" (Plate 175b by W. WRESZINSKJ 1923) represents a “red tiger-striped” in the original. This erythrism or flavism at that time, with its pronounced tiger stripes, had emerged from the F. ocreata markings. This process may be in accordance with the described decomposition or dissolution phenomena, thus even in present domestic cat lines this special wild colouring still occurs. It can, therefore, be assumed that similar things occur in other wild colours and markings and that, as a whole, the development of domesticated colours in cats is probably still in progress. Of the two cases of a sudden change to red in BROOKE’s "Nubian" strain, the Viennese cats represent the reinforcement of the tiger pattern, and the Dresden cats represent the beginning of solid colouration.
On the question of the breed-worthiness of the "Abyssinians" I would like to say that the clearly accurate restoration of the wild-type from the midst of domestically modified stock is rarely considered a goal in animal breeding. If consideration of valuable wild-types in setting up the German Longhair or short-haired tiger-striped appealed to me, this consideration did not guide me exclusively, and as far as it did it was about the structure and not about imitation. In the particular case of cat breeding, the treatment of the "Nubians" as breed cats is based on the concern for purebred breeding and preservation of two Shorthair breeds, which I shall discuss here, the thick-set short-haired tiger-striped, especially in the wild colour, and the domestic-coloured "slim breed" excluding the wild-coloured and patterned animals. From these two breeds comes a type which preserves remnants of markings on the wild colour and leans towards the slimmer shape. What made the "Abyssinian" popular in England and also attracted attention the continent was undoubtedly the catchy name that evokes the idea of the ancestral species and of ancient Egypt, and in general has the allure of exoticism, which has a special effect in cat ownership. At the same time, in this case of English Shorthair breeding, the fact that conformation cahracteristics are involved has inclined the breeder towards an African role model.
In the cases of the Blue Shorthair and the "Abyssinian" it has been shown that English short-haired breeds, in spite of a highly enhanced cult of colour combined with an emotional aversion to altering conformation, has occasionally been subjected to this principle. It has happened on various occasions, in each of which the conformation provided a natural and inevitable value. For Blue Shorthairs, Abyssinians and relatives, see SCHWANGART 1928/30, 9.
IV. THE TYPICAL DESIGN
Before I mark my own position on body shape in the classification of our Shorthair cat breeds, I will first turn to an element of colouration, the markings, and then the breeding biology and practical significance of colour characteristics themselves.
The two wild groups (F. ocreata, F. silvestris) apparently involved in the foundation of our domestic cat stock, as well as the candidate F. ornata, generally have the tiger pattern as the basis for their markings. It may be more or less blurred in adults, or it may be represented by rows of sports or an almost irregular mottled appearance. In the case of specimens in their first months of life, on the other hand, as far as my experience goes (descendants of F. ocreata, with a greatly reduced adult pattern), it is well developed. In adults it becomes more intense in F. silvestris than in ocreata (see Fig. 7 and 8), but it may also be more suppressed early on. Spotting appears to be the rule in F. ornata (Figure 9). According to VAN BEMMELEN (1931), there would be a primary state of mottling, and the banded markings would have developed from "stringing the patches together". In domestic cat embryos, K. TOLDT (1912) found the tiger pattern as a precursor of the domesticated pattern. Also VAN BEMMELEN considers the "transvere row arrangement to be the original skin pattern" of the mammals in general. But, as I said, this pattern originally consisted of rows of markings.
Complete disappearance of patterning is often encountered only in black or white domestic cats. An entirely unmarked "red" cat was described as a great rarity and depicted by H. C.BROOKE (1929c). Even with blue complete absence of markings is unusual. "Unmarked Isabels" [fawns] and "cream selfs" are ideals of English colour breeding that are hardly ever achieved. The Siamese has reached this stage, but in this breed you can allow a suggestion of stripes, for example on the legs, otherwise too many beautiful animals would be rejected.
The remnants of stripes in the house cat may be the basis of the tiger [mackerel] and the marble [classic tabby] pattern (see Fig. 1 in the text, pg. 98) can be briefly noted here, I will report in more detail about it later.
POCOCK (1907a) has emphatically pointed to the systematic and phylogenetic significance of patterns and markings, ahead of colour, and their merit in the classification of wild felids, and he rightly represented this same point of view as the basis for domestic cat breeds (1911). He added that such arguments could not dissuade the English breeding trend from disregarding markings. I have, as will be discussed further, when indentifying my breeds "Tiger-striped Shorthair" [mackerel tabby Shorthair] and "Marble Shorthair" [classic tabby Shorthair] combined features of form and pattern (1928b) and explained the status in more detail in my essay "Shape, Pattern, Colour" etc (1928/30, 4). In England, HC BROOKE (1929b) shared this opinion insofar as he acknowledged the breeding-biology significance of the patterns and energetically pushed for the dissolution of the English group of the "tabbies", the name for both patterns andu under which both were thrown together, and for the most part were restricted to shades of grey. On the other hand, this same author expressed strong doubts about the possibility of breeders ever becoming interested in Shorthairs based on pattern and form, except for his "Abyssinians." I am already justifiably pessimistic here, and this especially applies to the "grey" wild and slate-coloured "tiger patterns" to whose breeding conservation I attach particular importance, as will be shown. The disregard for these "ordinary gray ones" is especially entrenched in our country. The marbles, [classic tabbies] whose distinctive pattern attracts everyone's attention, have more prospects, and in addition to the red marbles, the red tiger-striped cats are also held in higher regard by reason of their colour. My prognosis in favour of a scientific future for cat breeding anticipates the old opinion, confirmed by my own experience, that in animal breeding perhaps we can hope for the establishment of a few exemplary breeding programmes through the interest of talented individuals.
No other pet retains, in such a high percentage, the markings of its ancestors. (The same can probably be said for the wild colouring, which I will discuss separately.) Equally peculiar to the domestic cat is the combination of patterns with various domestic background coloursin addition to the wild colour and related grey colour. We have patterned wild-colour, fawn, slate-gray, silver-colour, deep (chocolate) brown (simply called "brown"), "blue", "red"; the background colour can also change without affecting the pattern. Between these aforementioned shades, patterned domestic cats come in all sorts of transitional hues, although those such as actual "red" with its "dilutions" is set apart and has hardly any intermediate tones, and is often even found in white-spotted mixtures. Thus, the patterns, even the wild “tiger” pattern, are independent of colour, while the wild pattern is colour-dependent.
The fading of the pattern already established in the wild ancestors, and the state of its representation by a series of spots, or more rarely by the appearance of diffuse stippling ("Perlkatzen" [tipping]), also occurs in domestic cats. Transitional stages of surface colouring often appear under the white-spotted patterns between the patterned and solid areas (Tiger-striped-and-white, Marble-and-white). This type of spotting can also have a symmetrical arrangement, leading to the "masked" pattern. The other extreme is a condition which, compared to the wild-type pattern, reinforces the striping, and together with the tabby pattern creates the image of a "zebra-cat". I have seen such excessively sharp-striped specimens only in shades of grey (including the "blue"), whereas in the case of the "red tiger tabby" (as with the "red marble tabby") the contrast is weakened. It is feared that these varieties increasingly lose the intensity of their pattern in their mode of inheritance, which we must try to counteract, but as already mentioned, red very rarely appears as a solid colour.
The significant variability of stripe width and spacing, so that the number of stripes and the shape of the markings vary continuously from a long, clearly marked, oval-shaped stripe to small roundish dots.
An intensification of Wild striping - in this case only on the wild colours - is mentioned by ANTONIUS (1922) in regard to domestic horses. It is a type that appears as "mouse-grey and pale yellow in wild-coloured individuals, both in combination with a clear eel-stripe [stripe along the back], dark tint on the front of the legs, shoulder stripes and a greater or lesser degree of striping on the lower legs, hocks and on the sides the forehead. These striations are much stronger in some wild-coloured domestic horses than in their wild ancestors." Since the striped markings found in Wildcats (see above) are stronger when young, it seems natural to consider any strengthening of these markings in adult domesticated descendants as persistence of a juvenile characteristic. This occurrence is a phenomenon of domestication, well-known in breeding biology and especially visible in osteological examples, as already mentioned when comparing dog breeds (see HILZHEIMER 1931).
In any case, domestication of the cat has also had an effect on the pattern and colouration to opposite extremes: shrinkage and contrast of the striped pattern. Suppression occurs in the form of fading of the stripes, darkening of the background colours, breaking up of the coloured areas. The marble tabby pattern, as I will show, is equally effective in increasing the area occupied by the coloured bands. VAN BEMMELEN (1931) gives an accurate example of the process of fading in post-embryonic life. In its youth, a “black” cat was actually still patterend with superficial markings and all traces of pattern disappeared after its first year of life. Such findings are in addition to K. TOLDT's findings that pattern is preceded in the embryos by surface flecking.
In the "tiger" as in the "marble pattern" of the domestic cat asymmetries occur. These are less conspicuous in the former than in the latter, but this does not indicate varying frequency. The marble pattern simply makes them stand out much more. Such asymmetries are also common elsewhere. Thus, K.M. SCHNEIDER (1930b), presented a case that came to his attention, depicting a young eland from both sides. O. ANTONIUS (1930) reported the same thing about a zebroid (mountain zebra mare, Malay pony stallion), where the asymmetrical stripes of both sides of the body was particularly pronounced on the neck, in such a way that the number of stripes on both sides was approximately equal, but the "pattern of striations is quite different". In SCHNEIDER's case, conversely, the number of stripes differed. On our domestic tiger-striped cats we have both types of asymmetry: the pattern of stripes probably differs in the vast majority of cases, and the number of stripes quite often. Also in some other species these asymmetries appear quite common, such as, according to ANTONIUS (1931a) in the true Quaggas. I come to the same conclusions that V. HAECKER (1918) and E. ESSKÜCHEN (1929) drew from such incidents after reviewing the marble tabby pattern of the cat [See Footnote]).
[Footnote: A particular type of asymmetry is that of VAN BEMMELEN (1931), especially for the tiger's stripes (Uncia tigris, Fig. 4) and the leopard’s spots (Fig. 3) and Jaguar demonstrated at the meeting of the both striped or spotted sides along the midline of the back; they are "not exactly symmetrical," but "interlock into one another" where they meet. Based on this, the author hypothesises on the origin of the vertical rows and then on the horizontal striping in mammals from this overlapping variegated coat. In the case of the domestic cat, the conclusion on this particular type of asymmetry is made more difficult due to the often unevenly curved stripes and, respectively, their number and course. Since the publication of VAN BEMMELEN's work, there has not been time for appropriate investigation.]
I have already touched upon the important question of breed-biology of a constitutional, eugenic superiority of the wild-type pattern persisting in domestic animals. I spoke first of the wild-coloured and other grey "tiger cats". In the domestic cat the wild “tiger-stripe” pattern is often associated with wild colour and its related grey, as well as with other colours, including the telltale "diluted" degenerate ground colours from pet breeding, and the wild colour can be associated with the non-wild, separately inherited "marble pattern." Therefore, here is a separate examination of wild pattern and wild colour, considering their merits, with associated questions such as the effect of combining the wil pattern with "diluted" colours and the domesticated intensification of patterns. In addition, the factor of body shape plays a part, howeever I must postpone the treatment of that topic. First of all, investigations are complicated by the high degree of cats persecuted and maltreated by physically and mentally worn-down cats, giving more reasons why it is necessary to restore the domestic cat as a valued home and farm animal.
The science of inheritance reveals a contrast between "striped" in the strict sense (broad bands) and "lined" (narrow bands), where a dominance ratio applies (see BAMBER 1927). Among these factors,the distribution factor for wild colour (in the tiger-striped as well as the marbled) known as "ticking" - "agouti" in WHITTING (1918 after BAMBER and 1919) - occurs in these experiments; it dominates over non-wild colour and should again show a dominance ratio, an issue where we lack the required clarity. As a rule, in these experiments, stripes and wild colour were treated as the available combinations and no mention was made of different coloured stripes (BAMBER 1.c.). I will occasionally refer to these results when discussing the marble pattern.
Next to the tiger pattern, as already stated, there is a second pattern in domestic cats, which we call the marble pattern (according to POCOCK, "marbleds" is termed "blotcheds"). Pocock (1907a and 1911) drew attention to the important circumstance that with this duality sets the domestic cat apart from other felids. He sought to explain this by assuming an unknown extinct wild species which was marble-patterned, and according to this domestic cats were not only a race, but to some extent were different from the others. I was unable to agree with his opinion (see SCHWANGART 1928, 1929), but instead consider the marble pattern to be a domesticated mutant. Under this assumption, hardly any case is known where the domestic life-style has made such a dramatic effect. It appears a violation of a law valid for all the wild realtives of this pet. The marble pattern comes, as I have said, in Longhair as well as Shorthair, however, as I will show, it is mostly only the latter type that is used for breeding. I have already mentioned that bearers of this pattern occur in the same colours as the "tiger cats". Therefore, it is most probable that even in cases where a pattern is so greatly obscured by domesticated colour changes that its species can no longer be recognized from the remaining traces, various colours have gone along with the marble pattern as well as with the tiger pattern. For example, in the case of "smoke cats" (where the basal part of the hair is white to white-gray, and the terminal part is a smoky black-brown) one can often still distinguish the “smoke tiger” from the “smoke marble” by the tinted areas. From this we can assume that in embryonic development the marble pattern also precedes the solid colours, as TOLDT (1912) has shown for the tiger pattern. Data from embryological investigations could also contribute to the genetic relationship between the two patterns, and to the question of how the marble pattern emerges. It goes into the particulars of the parallel behaviour of both patterns towards the colours.
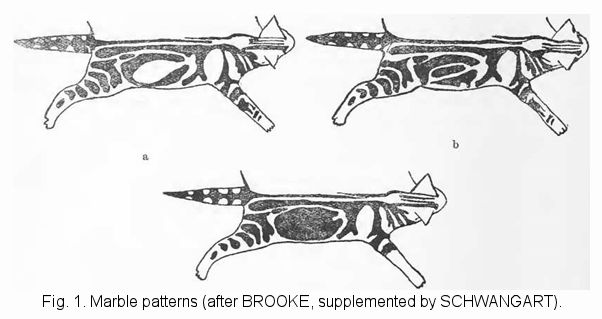
Differences in the behavior of the two patterns are found in the rarity of the marble pattern to disintegrate into smaller spots, its less frequent tendency for markings to become narrow beyond a norm, and its opposite tendency for the markings to broaden, which can lead to the establishment of solid areas. I will continue to report on cases where the broadening of markings are recognised straight away when discussing the various modifications of this pattern.
The marble pattern consists of exceedingly beautiful "baroque" [ornate] markings (see Fig. 1), within which there is a particularly noticeable elliptical curve that bends around and almost closes ("spiral", "horseshoe-pattern", "wheel"). The official name "marble pattern" can, therefore, only be considered in contrast to the "tiger pattern". This pattern is subject to a myriad of hard-to-analyse variants. An exhaustive classification system, even if it only includes the breed's "permissible" patterns, rather than deviations upsetting the pattern’s harmony has not yet been set up. The patterns in Fig. 1 illustrate only the basic principles of such modifications. Firstly, the inner "wheel" can either be “filled” or "not filled" with the background colour (a) or contain a coloured internal spot (b) or filled in the same colour as the markings (c). Secondly, besides the most prevalent interrupted cranial markings (a, b, and c), it may have a second broad ventral marking illustrated by the filled wheel (c). If one disregards the "basic scheme", then the frequent occurrence of further open spaces can be determined, which lead to the many complex irregularities in the marble pattern.
On the "marble" a "triple dorsal line" is desirable, while most "tiger striped" skins carry one uniform stripe [see note]). Isolated cases with two, or even three, distinct dorsal stripes occur, however, and I have also seen separate dorsal stripes on skins of F. silvestris (Zoologial Museum, Berlin), which means that this formation should not be considered as exclusive even in the "species description" of marble according to POCOCK. Noteworthy in this respect are also some details in OGNEV (1 c.) regarding the dorsal markings of Russian wildcats. For F. silvestris silvestris SCHREB says OGNEV noted the individual occurrence of "two black stripes along the back." In his description of F. ornata caudata GRAY states that this species has normal "undifferentiated longitudinal stripes" on the back, they were replaced by points indicating emerging row marks (which corresponds to the overall pattern of this cat), but as individual differences he often found "three rows of spots indicating weak stripes" and in a particularly strongly marked specimen "indications of two to three dorals stripes, interrupted here and there."
[Note: In the section on marking patterns, the terms "tiger" and "marble" also include individuals with those patterns, which, according to their body shapes, do not correspond to the breeds of shorthair named later by me as "tigers" and "marbles". It depends on the patterns first.]
The oldest known description of a marble pattern to date is that of LlNNAEUS (1759), who based his Felis catus on this type, in the erroneous opinion of having a Nordic wild cat in front of him. I have followed POCOCK'S nomenclature. According to this, the name F. silvestris SCHREB also applies to this wildcat in the current wide term. According to POCOCK's opinion of the independent types of domestic cat carrying the marble pattern (both short hair and long hair together!) he gave the species name F. catus fell to them - according to my opinion, which does not recognise their independent style, this name is obsolete. In any case, there is an interest in the question of how far back the marble pattern can be traced. After all, it can be stated that it appeared long before LlNNAEUS's description. Here I present two proofs. Unmistakably, a complete marble pattern appears on KARL RUTHART's large group picture "Odysseus at Circe (1666)" (Dresden State Gallery, Fig. 16), and from much earlier I found this distinctive pattern shown in the Old Pauline (Great Greek [i.e. coastal areas of Southern Italy]) representation of Fig. 17 (after O. KELLER 1908), dating from about the 4th century BC. Assuming it is true to nature, which is supported by a current parallel case, this pattern appears here as broken rows. Even the moderate performer has not escaped the special grouping. Accordingly, the marble pattern goes back to the time of the ancient Greek (and the ancient Egyptian) cat husbandry, in this case probably because it derives from the strain F. ocreata strain. For Longhairs it has developed on the F. silvestris conformation.
Just as POCOCK, in his propensity to label the wearers of this pattern as a particular species, regards the triple dorsal stripe as fully characteristic of marble, he denies the existence of mixed or intermediate patterns between tiger and marble. I have come across both kinds of combinations, the first often blends the two in different variants such as the one shown in Fig. 18. The only observed case of actual transition between the two Fig. 19 (a and b) deserves attention as a comparison of both sides again shows asymmetry [See note]) In addition, the bands break down into spots on the ancient representation in Fig. 17.
[Note: The case of a one-sided circular arrangement of spotted lines is seen on a young specimen of African F. ocreata, pelt in the collection of the Berlin Zoological Museum. The spiral direction was contrary to the norm in the marble domestic cat. Of course, I also learned of specimens with the "wheel" pattern reversed, but in those cases the deviation appears to be secondary to the breaking up of the markings. Thus, the Berlin case may well be a proof of the tendency of the circle pattern within the tiger pattern even in the wild, but not in sucha a way tht it imitates the marble pattern.]
From the still incomplete crossover results between bearers of the tiger pattern and of the marble pattern, one can conclude the Mendelian mode of inheritance. (I will later come back to the experiments as a whole.) Intermediate forms as a result of crossing (here as "intermediate" heterozygotes) would then be excluded and the two individuals shown in my pictures would require a different interpretation. I myself know of cases of cross-breeding where poorly-marked marble patterns appeared in addition to correctly marked patterns of both parents.
If necessary, cases such as that in Fig. 18, which so strongly give the impression of being a mixture, can be interpreted as "emerging marble patterns." Obviously, the transition process from the traditional tiger to the domesticated marble pattern is shown in Fig. 19. To be understood as stages in the process of change, these pictures signify an early stage in the development of the marble pattern, with the typical "horseshoe" already present and developing asymmetrically.
Thirdly, except for interpreting the intermediates as hybrids as a result of crossing or as transitional phases, there remains the "anomalous" mutation. With regard to the two types in question, that shown in Fig. 18 and that in Fig. 19, I will come back to this possible interpretation.
If, like myself, one considers the situation shown in Fig. 19 as a genealogical clue, as evidence of transformation, then the initial conclusion is that the process is gradual. All sorts of additional conceptual possibilities remain.
The transformation could take place in the developed animal or embryo, both modes account for other changes in the marking pattern and the domestic colouration, or there were types of change – both in the finished pattern or steps toward it – appearing in the permanent state in the embryo, whereby the step-by-step process would be limited to the germinal elements [the gametes]. Triggering of this process by domestication is conceivable as somatogenic or plastogenic. A progression through several individual stages to the finished pattern is conceivable, or step-wise over generations, with unequal final stages, or an already complete pattern; vis-a-vis the idea of persistent degree variations.
Settlement of this problem would require a knowledge of the origin and early development of the animal of Fig. 19, further observations of it and trial breeding with it. The animal arrived at a cats’ home already pregnant, died soon after and thus lost the embryos it carried. Nothing useful could be found out about its ancestry or inheritance [See Note].
[Schwangart writes “ancestry or preformation.” Preformation is an early development theory whereby an organism develops by unfolding preformed parts; in the current context, “inheritance” better fits the context.]
The conceptual possibilities of the transformation process already mentioned for this case are also applicable to the categorisation of Fig. 18, if one also wants to attribute such images to a transitional event. In this case the possibility of an erratic process next to a stepwise one could be considered.
Assuming that the process of change represented by Fig. 19 still occurs now, which we cannot dismiss, then it must be regarded as exceptional these days, unless the possible instances of an additional occurrences of the new formation in the finished state (i.e. the complete marble pattern) is the rule. In the event that the frequent intermediate stages of the type of Fig. 18 also represent transitions, this type of active transformation is reasonable.
In every case, under domestication, the process of change would be driven by the environment, including changes in the marking patterns and the domesticated colouring. However, while alterations such as albinism, melanism, "dilution of colour", fading and spreading of pattern and colour, environmental stimuli, by altering the constitution, also affect the qualitative effects of these trends, for example the specific form of the marble pattern is caused by mutation and does not depend on the environment at all. If the environment triggers its appearance, it must be "orthogenetically" predetermined [Orthogenetic: an innate tendency to evolve in a definite direction] [See Note]).
[Note: In a compilation of "References to Some Italian Works," O. DE BEAUX (1929, pg. 56), casually mentions in his "Remarks on African Cheetahs," "the coexistence of a longitudinally striped, clouded leopard-like" type alongside the striped wildcat type in domestic cats and assumes a particular distribution of this type, which, in spite of the somewhat incorrect comparison with the clouded leopard, can only be understood to be the marble pattern, "in cities" where "these animals . . . often have no master and are not subject to the slightest breeding attempts on the part of man." This is a conjecture about the type of environmental influences that produced the marble pattern. It would be a domesticative milieu, but one in which the animals were not actively nurtured. This gave the marble pattern its specific character that was also related to constitution. A more independent way of life, with a certain number of domestic influences coming into effect, meant that the animals would not become solid coloured, but would remained patterned, rearranging the existing wild markings. — However, I have not found DE BEAUX’s assumption of a more independent existence upheld in either in our reagion or in others I know to be associated with a greater frequency of the marble pattern. In several areas, this pattern is favoured by the common people, so that cats with this pattern are rather well cared for. The Shorthair breed is currently maintained by so few that in most places, we can say that matings are "not subject the slightest breeding attempts".]
The mode of transformation, reflected in Fig. 19 (a and b), of changing the course of a line, is fundamentally different from other kinds of domesticated colour change, including the suppression of markings, thus in this case we see cases such as Fig. 18 where images correspond to a conversion process. Assuming they do, then the condition of Fig. 18 shows the process of one pattern degenerating in favour of the other.
It is improbable that a transformation in the manner of Fig. 19, which implies the stripes both reduce in number and divide into sections, shold take place post-embryonally (e.g. in the "postembryonic progressive phase" BR. EGGERTS 1931) whereas a transformation in postembryonic life would be plausible for the process suggested by Fig. 18.
On the whole, one is reluctant to accept two multiply contradictory processes, such as that according to Fig. 18 and that according to Fig. 19, as paths to the same goal. And even the interpretations, which are decidedly more appropriate to the picture itself, are different for Fig. 18 and related phenomena than for the case of Fig. 19.
As previously mentioned, possible interpretations other than transitional forms and intermediates due to hybrids can be considered mutative abnormalities. In the case of Fig. 19, however, it is hard to imagine that the almost obvious type of rearrangement of the pattern would not correspond to a normative principle. Interpretating it as an abnormality or a blended product does not suffice for this picture. For cases according to Fig. 18, on the other hand, the last two interpretations appear to be possible options. The most convenient of those is the mixed or blended form, but this is contrary to the previously mentioned experimental results and to our experiences with crossing, which of course need further investigation. When interpreted as an anomaly, one first tends to think of elements of the ancestral pattern persisting in a haphazard mixture with those of the newly emerging pattern. In principle, an anomaly arising in this way is contrary to the normal method if, like me, you believe you recognize this type in Fig. 19. Such an exception is unlikely.
One solution is to assume that the anomaly occurs due to the re-appearance of the wild pattern with signs of the domesticated pattern breaking up.
In the view that such a process of "reversion" is happening here, I am encouraged by the not uncommon cases shown in drawings a, b and c in Fig. 2 (pg 107). In each one we see an irregular marble pattern within the area of the "spiral" (wheel, horseshoe). The drawings are true to photographs. The pictures should not yet be considered stages, but simply as individual cases. The overall shape and details of the part of the pattern depicted are influenced by the respective position of the animal when photos were taken. This position is characterized by an indication of the abdominal and the line of the back. Fig. 2a shows the portion of the pattern representing an "internal spot" of the "wheel" dorsoventrally split, with a tendency to form coarse stripes. 2b shows finer stripes in the same place, dorsally connected together, running into ventrally oriented points (as true tiger stripes can do). 2c has a uniform "internal spot", that expands ventrally into short stripes. If one compares this latter picture with Fig. 18, thus in this case of blending the patterns the ventral end of the "inner spot" is combined with with stripes and we can distinguish a weak streaky offshoot and stronger narrow stripes crossing the inner spot. Environmental influences could be directing this reccurrence of the wild pattern on this and other domestic cat ground-colours. According to the pictures, it is probable that this would usually or always be a partial new formation. In a marble [classic tabby] cat presented to me the initially correct pattern had been disturbed by the strong appearance of horizontal stripes. The striping lies outside the "wheel"; the process as such, however, is similar to that presupposed here, and follows the process discussed in the next paragraph.
I have observed isolated cases of changes in the marble pattern during postembryonic life, including one in which the value of the animal was impaired. The observed changes were the fading of the pattern and the broadening of the stripes, but never extended to modifications inthe course of the stripes. See also the special case at the end of the previous paragraph, which mentioned an increased appearance of the other pattern. These took place over the course of several years and generally remained small. The marble-breeder Dr. Med. HEINE-Nerchau has observed changes during the first months of life, consisting of the appearance of lighter hairs in the darker pattern itself. It was always wild-colour hair that appeared. "Beautifully marked" silver marbles lost considerably in value as a result.
According to WHITING’s crossing experiments (1918, 1919) mentioned in the earlier discussion of the tiger pattern, of which only the second mentioned in the original is known to me, and BAMBER’s abstract (1927), the "striped" and "lined" tiger patterns (an explanation of these terms appear above) are strictly separate in inheritance ("segregate sharply"); the "lined" pattern is dom,inant to the "striped" pattern. Both variants of tiger pattern were dominant over the marble pattern ("blotched"). However, the allele character of the three patterns is not considered assured. In general, this means "inheritance not fully worked out". Mrs. BAMBER rightly writes of WHITING‘s attempts to be factually inadequate, which were partly due to material difficulties, and that his presentation of meant the English person found it "very difficult to follow his remarks". Contradicting the original tables, BAMBER considers the stronger patterns, "blotched tabbies" as "striped tabbies", only when occurring in combination with wild colour ("ticking", "agouti"): “Ticking or agouti in combination with banding gives the tabby pattern.” The tables themselves include "blue, yellow, cream stripeds" and just "blotcheds" and even "tortoiseshell stripeds" and "blotcheds". As previously mentioned, so far I have not seen a significant narrowing of the norm in the marble pattern, the fact that the "lined" factor is reserved for the tiger pattern as opposed to the blotched factor, as with WHITING and BAMBER, appears valid. In contrast, the more common form of this pattern, wherein for example, ften involving the completion of the "horse shoe" (see Fig. 1 c) is often ignored, as has the factor for the composition of spotted rows which is so common in the tiger pattern ("spotted tabby," which BROOKE 1929 b simply calls "tabby"). — The absence of "ticking" leaves the banded markings "present", but only as shadows ("ghost markings"). I have already considered the gradual grading of the "ticking or agouti" when reviewing the tiger pattern, as well as several times regarding the occurrence of domesticated colours in wild-patterned domestic cats; but these the stripes can stand out even more sharply than is normal in the wild colour. Further, I recall the fact that all the pairs of features, even the basic plans of the tiger and marble patterns (Figure 19), are more-or-less opposite to each other up until the epproximate point of transition from one to the other. Nowhere does WHITING‘s cross-breeding table show intermediate heterozygosity in terms of opposites; Mendelian inheritance appears throughout.
In reviewing the tiger pattern, I touched upon the important question of the constitutional (eugenic) advantage of wild-pattern domestic aimals, where the domestic cat has a preferential position, and I postponed this discussion due to interaction of this factor with wild and domesticated colours, peculiarities of form, and because of the related competition between the marble pattern and wild pattern.
The marble pattern presents (in my already well-founded view) not a wild pattern, but is something fundamentally different, it is a natural precursor of the solid colour of the domestic cat. As far as my particular marble breed is concerned, it should not be combined with a wild-type conformation as that is often already connected to the tiger pattern. The marble pattern goes together with the wild colour and related grey colour as well as with the domesticated colours.
As regards the eugenic question, I think it noteworthy that complaints have never been made about "red" and pale yellow tiger-like marbles, as well as selected Isabelline and cream cats (more about these in the sections on colour) that are heavily patterned on the trunk, and an omission by the marble-tabby breeder Dr. Med. HEINE, who prefers silver marble, that domestic cats in colours only distantly related to the wild colour (discussed further in the sections on colour) are preferred for their special beauty. He wrote to me "straight tigers and marbles are characterized by special intelligence," and with regard to the character of the marble patterned cat, he explained that the majority of them are calm and steady in their behaviour (See note). I have noticed remarkably steady, composed behaviour in the silver marbles as well as in the wild colour, during transport by train or car, even in a sack, which is dangerous or even lethal to them if you do not let their head peep out. However, we doubt that such judgments apply to every single well-kept cat. The mental state cannot be unalterably linked to a factor such as pattern, or indeed exclusively determined by form, pattern and colour together.
(Note: Since this benefit is combined here with the wild pattern, I accept the view which is common among scientists, of the absolutely "stultifying" effect of domestication. While this may necessarily depress some faculties, including intellectual ones, it can develop others that are significant to us, to a varying degree, depending on the role we assign to the animal. Incidentally, in the special case treated here, besides the tiger-pattern, the domesticated marble pattern must be considered.)
The pure breeding of the basic marble pattern appears to be easy to achieve according to previous experience. This matches the assumption it is receessive to the tiger pattern and to several solid colours. The difficulty in breeding this pattern is due to the strong preponderance of cases where the pattern is imperfect (aberrant). The first requirement is to try to obtain raw material that is a correct as possible.
By virtue of their diverging directions of development and the variety of their modifications, the marking patterns seem to me to be a particularly promising case for researching causal relationships in the origin of mammalian, and indeed general, animal markings. A good overview of works in this direction up to 1929 is given by Edm ESSKÜCHEN. For more details, refer to his composition. V. HAECKER (1918) drew attention to the various explanations for the variability in the number of transverse bands, the significant individual differences, the asymmetries, and all the facts set out in my description of the tiger and marble patterns in domestic cats. From the currently state of knowledge of the independence of laws that impact mammalian skin-markings, HAECKER concluded there was metamerism (note: metamerism is the phenomenon of having a linear series of body segments fundamentally similar in structure, but not all identical in function, in mammals often manifested in skin pattern). On the other hand, VAN BEMMELEN (1931) returns to a consensus of both because the number and arrangement of transverse rows often matches the number of vertebrae. These can be increased "by doubling the stripes" or can be diminished "by variable degeneration". The layout of the domestic cat’s marble pattern makes this a type that obviously defies metamerism. Even with the occurrence of a transitional state between tiger and marble patterns, as in my Fig. 19 and my interpretation, if this was how the marble pattern arose from the tiger pattern, the metameric nature of the tiger-pattern gives me no reason to assign the marble pattern to metamerism. The change of the course of the line to a completely different system illustrates the intervention of a different operating principle independent of metamerism. The same applies to the establishment of the domesticated piebald pattern as the successor of both patterns. Determining factors, developmental facts and alternatives do not yet provide the information required for investigating these wide variations of pattern, symmetrical, asymmetrical, (see note).
(Note: Only a small measure and a small scope of inheritable and uninheritable asymmetries correspond to the general state of the organisms. If one thinks of these shapes as being dependent on certain nerve effects, which will be discussed later, these actions will not be symmetrical even if they strive for symmetry.)
ESSKÜCHEN (1c) presents a grouping of mammalian pattern types, namely, "tiger-stripe, tiger-spot, dappled, roan" opposite the "metameroid" ("piebald, patched"), where both groups were "unrelated" to each other. VAN BEMMELEN (1 c.) saw, as already mentioned, a phylogenetically common initial state in the spotted lines, this applied to mammals in general as well as to his main subject, the cat. He sees in this hypothetical initial state of all drawing, and moreover of spotting and monochrome, "the image of a dress of overlapping scales," and investigated the origin of the blotched rows as "an accumulation of pigment in the groups of hair behind the scales and in the tail-end of the scales themselves. During the degeneration of the scales, the pigment was transferred to the hair that formed in their place". HAECKER sought the cause of the total differentiation in a distinctive rhythm of skin growth, he also demonstrated a certain type of rhythm (checkerboard type) in vertebrate embryos (Axolotl) and found that the main divisions zones it created contradicted the creature’s segmentation. "In the Axolotl a causal relationship between transverse and longitudinal growth lines on the one hand and the transverse or longitudinal striping on the other hand can therefore be demonstrated" (ESSKÜCHEN). HAECKER concluded that the markings of our domestic mammals were based on the same process that he found on the axolotl. Undoubtedly, the image of the two marking patterns in the domestic cat, and more importantly that of a genetic transition between the two, as in my Figs. 19a and b, can only strengthen this assumption of such a fundamental basis.
W. JANKOWSKY (1931) sought to track down the underlying and contributing factors behind the puzzling variety of markings. In his "Investigations into Hair Colours" he emphasizesdthe "undoubted influence of the nervous system, which manifests itself in the often symmetrical coat markings" and "still requires complete research". The patterns and the "metameroid" spotting are summarized in the sense of a symmetrical coat drawing. It has already been mentioned several times that asymmetry must also be considered both individually and in connection with symmetrical basic features. VAN BEMMELEN, even with regard to the spotted lines which he regards as primary, grants the nervous system a certain influence, inasmuch as "Pigment accumulated where sense organs were located and in many cases asserted themselves in their original sites when the organs differentiated or degenerated." W. JANKOWSKY means central nervous system activity.
We have to agree with ESSKÜCHEN’s conclusion two years ago, that HAECKER’s discovery of the rhythm of growth had created a "new fundamental," but nevertheless "we are still very much in the dark," and we maintain this judgment even after the recent further studies where one of the investigators, JANKOWSKY, has spoken out (see above). The main question now probably concerns the factors that cause the nervous system to b e active in directing certain systems of markings, and in the case of the domestic cat even in alternating between two fundamentally different marking patterns that exist side-by-side and where the new one apparently replaces the old transitional one. It is hardly wrong to look for the triggers of the nervous system‘s directing action in the area of the constitution, which is evidently already effective in changing colour. The question of metamerism with regard to the pattern types is, therefore, not obsolete, nor can other determinants be excluded. Rules of correspondence of certain constitutional states (physical, mental constitution) with certain types of pattern will be uncovered by careful observation of the animals; to find out how the nervous system then trigger certain patterns, and to solve the question of why the constitution is manifested in the type of pattern, we must first have to look for the mechanism.

ESSKÜCHEN considers "undoubtedly" that the difficulties in solving the relevant problems are "much greater in domestic mammals" than elsewhere, "because the causal complex is probably much larger". To some extent this will be true, but we must not overlook the fact that the greater variety and especially the existing variability of domestic mammals also provides a strong advantage, which can be clearly demonstrated by the example of the domestic cat. In general, for the whole problem, I would like to re-emphasize the probability of complex causal factors.
V. COLOURING (OF HAIR, SKIN, EYES ETC.)
In an overview of data on domestic cat colours, excluding the markings already discussed, it is best to begin with the wild colouring, which has played a role here in other contexts i.e. the wild species, Abyssinian breed, markings. A wide range of variants were already recognizable in the wild material, likewise that it can persist with the wild-type striping and marble patterns between different coloured areas, sharply defined or transitionally, in piebalds.
I have not attempted an analysis of "wild colour" gradations in the domestic cat. The overall impression corresponds to the picture reproduced by HAECKER (1918) and ESSKÜCHEN (1929), according to whom it generally results in a "belt-like or zonal" pigment arrangement, wresulting in numerous gradations, such as greyish-yellow, greyish-brown, reddish-brown, etc. I found the reddish-brown already emphasized in a Sardinian island form of wildcat (SCHWANGART 1928b, 1929a), with their fox-like appearance, with contrasting black, rather strongly developed ear-tufts (specimen in the Berlin Zoological Museum).
Regarding the colour difference of the hair on the basis of different pigments or only on the different density of the pigment arrangement, HAECKER took the viewpoint that in the wild-colour several types of pigment would be present, by adding to the wild-grey colour "colour variants where wild-grey pigment distribution in the individual hairs may still be hinted at, although one or two types of pigment found in the wild-type are missing." W. JANKOWSKY's (1.c.) new investigations, by which the pigment problems that had reached "a dead end" were for the first time chemically and experimentally tackled and are apparently closer to being answered. These investigations on the question of unity or multiplicity of pigments showed that "the natural hair colours are merely different oxidation states of a chemically undetermined basic pigment substance", therefore "racial colour differences are not just quantitatively based on "the amountof the same pigment", but are based on the qualitative "chemically distinct pigments. The type of zonal ringing of wild-coloured hair was specifically found by in rabbits, guinea pigs and wild boars.
That there is strong variation in the range of wild colouring in domestic cats has been previously mentioned; for example it is seen in the presentation of brown to smoky blackish hair with a white or whitish basal region. Rarely, there is a kind of reversal of this : white or light yellow-gray hair with deep iron-gray basal reagion.
ESSKÜCHEN states that, due to its rich material, pets generally show a primacy of the wild colour in their constitution. With, or after, the wild colour, the most favoured variants are "red" (not yellow!), Reddish brown, brown, grey-brown, grey, with preference for erythrism, which is "still close" to the wild colour in that it contains reddish brown tones "but in addition to these come black-brown and yellowish- to grey-white". Regarding the domestic cat, we would have to say that the pronounced "red" tones in the wild colour regress more.
In any case, ESSKUCHEN's results, drawn from much experience in favour of the constitutional primacy of the wild colour and related colours, support my efforts to preserve and improve the domestic cat; fortunately they still present strong possibilities for breeding the wild colours. Unfortunately, there is a prejudice against "grey" cats ("peasant cats"), which are the main bearers of this beautiful pattern, which is further fuelled by backwards literature. Thus in an area generally not hostile to cats, bird protection guides may accuse them of having a predilection for catching birds. The author attributes this to confusion: he has not observed this trait to be tied to colouring, yet contempt for these cats makes them more neglected than others. My experiences, especially with the sturdily built wild-colour cats testifies the opposite, that these stocky cats are less likely to hunt birds.
Separate testing of wild pattern and wild colour for their constitutional value thus makes it easier to arrive at an positive result in terms of colour. The same applies to the corresponding comparison of the wild colour with the pattern (in general, including the tabbies). If we could assume there were indications of desirable qualities in cats with the wild and marble patterns, then the combination of these patterns with domesticated colours, specifically with the "diluted" colours in future requires closer and more detailed observation than has been done so far. The continuity of the original pattern or both in "diluted" colour could e.g. For example, indicate a constitutional restriction limited to certain features. In addition, it will appear that, in this respect, the "dilutions" also rank differently.
In addition, ESSKÜCHEN considers the silver grey, which plays a main role in marbled domestic cats and is also associated with the tiger pattern, closely related to the wild colour. He regrets the decline caused by inefficient breeding of the "silver-coloured" Dutch cattle breeds which were characterized by solid constitution.
To compare this colour variety to the "wild colour" we must consider the wild coat. There are enough mammal species whose natural colour, in accordance with the aforementioned viable concept, should not be treated as "wild colour" but as "diluted". In the case of the domestic cat, the silver tint, which is here connected to intermediate shades of slate gray and blue, is also found mixed with the wild colour (the long hair inside the ears being pure silver) and in the mottling of some F. Ocreata; this silver component is found in the "Abyssinian" domestic cat. Furthermore, in favour of the relation between colour and pattern, and the tendency of the silver colour to associate with markings, I can say: I have barely seen unpatterned, truly silver-coloured cats. In colour, the silver tone is close to a rather rare pale, often yellowish light grey. In certain cases, however, it is in direct contact with white: on the chin of the silver marble and silver tiger, a place where white is not permitted, this transition often causes difficulties for the judge. With the chinchilla colour (gray-toned hair with black tips, selectively bred in the Longhair), care is needed, according to "Domestic and Fancy Cats", to prevent the desired silver grey colour turning into "dirty white". With its propensity for a crimson nose leather and darker skin, its patterned silver tones are close to the other shades of grey. OGNEV (1.c.) describes extremely grey, we would call it "slate-gray," specimens of Felis ornata, a colour found in many domestic tiger- and marble-tabby cats.
As to the question of the reason for constitutional advantages of wild-coloured and closely related domestic animals - resilience of all kinds, breeding ability, type of temperament - ESSKÜCHEN develops the idea of a physiological effectiveness of the characteristic itself, by virtue of the inherent nature of the skin. This is also the case in the internal responses of these animals, for whom he also gives examples of particularly favourable development. For his reasoning refer to his work. One may well agree with it, while also assuming a second connection, that of the immediate merging of a favourable constitution with the characteristic, whereas the domesticated colour change, along with the loss of markings, altered constitution, in some cases possibly degeneration, may also signify their disruption. By the way, ESSKÜCHEN himself cites cases where this opinion applies. The same connexion is confirmed by the results of W. JANKOWSKY (1.c.) on human hair, where "in addition to its general significance for almost all metabolic processes, oxidation also plays an important role in the development of hair pigments." The degree of contrast in the cat’s markings could also be a sign of changes in constitution (the case of the "zebra-cat").— In all of this, it should be reiterated that animals naturally have different colours in their natural state, andthis also implies various implications for the constitution. The zebra-cat is modified from the wild stock, the zebra itself normally has this intensity of pattern, the wild lemming can naturally be tortoiseshell. The African Wild Dog (Lycaon) is normally seen in the colours of the "Spanish" cat, the lion is a "diluted colour," and the polar bear is normally leucistic. However, even for wild animals, the possibile significance of the colour as a feature of constitutional (in this case species-related) peculiarity (physiological, manifestation of character) cannot be rejected out of hand.
Black is known to be one of the most common colour variations even in wild felids. For examples, see SCHWANGART (1929, a). Melanism is further shown by Felis daemon SATUNINS (1904), S.J. ÜGNEV (1. c.) admits to the possibility of a Silvestris house-cat crossbreeding for these animals, but tends to designate them as wildcats and he considers a form of geographically limited melanism to be feasible based on the frequency of this phenomenon in wild felids. As a counterpart to the well-known breeder‘s rule, which applies to various pets, and apparently also for the domestic cat, "black calls the white", I also know the opposite process. A show-winning white animal later had a disturbing black spot on its head. In general, this case is evidence of colour conversion in post-embryonic life in the domestic cat, which I have previously discussed with examples.
Seasonally, according to the complaints of several cat owners, some, but not all, deep black cats turn slightly "rusty". There is also a persistent black brown with a "foxy" tint. On the other hand, between these deep browns and the deep orange to light red-brown of the "reds", which are the transitions towartds Flavism, there seems to be a gap in the colour scale. In any event, "diluted" black does not always give blue, there could be two kinds of black. According to JANKOWSKY (1. c.), "Rutilism" in human hair is closely related to black, insofar as red-hairedness comes about by darkening (lower oxidation) and blondness comes about by "brightening" (stronger oxidation) of black hair.
ESSKÜCHEN mentions the view of ADAMETZ (1904) and TELSCHOW (1911), according to whom melanism is a frequent consequence of strong inbreeding and where it is simply a result of domestication it indicates a weakening of the body through structural degeneration, because the cells serving the dye regulation can no longer do their job. The role attributed to inbreeding is out of the question for the domestic cat, which is very prone to melanism. Selective breeding of Shorthairs has barely started, and in the breeding of Longhairs, black is one of the neglected colours. ESSKÜCHEN also cites the widespread favourable assessment of black, according to which melanism is considered a "consequence of a kind of re-creation of constitutionally weakened animals", because, for example, "cases of severe pigmentation often occur in crossbreds of low-pigmented breeds." I have never heard any complaints about poor constitution in black cats.
What is noticeable in the domestic cat, is the tendency of black cats to have sleek shapes. So far, I have seen more Black Shorthair cats, which meet the requirements of my slender race, than in all other colour varieties together. Black was followed in this by blue (see "Blue Shorthair"). If we interpret the features of my slim race as "degenerative," then black colour would be one of the first indicators. Incidentally, every cat show and every local cat population contains its fair share of heavy and medium-weight Black Shorthair cats. Under Longhair, I think that only black has a regular tendency towards a slender shape (overall, more sharp-pointed) acceptable, but it is much less of a tendency than in Shorthairs. The danger of spreading sharp-pointed shapes and slender forms in the long hair appears due to the cheap prices of such exceptional raw material and possibly to the haphazard production of half-Angoras by mating Longhairs and Shorthairs together.
"Acromelanism" is considered an initial state of blackening (snout, ears, feet, tail tip symmetrical black), a classic example of this being the Russian rabbit. This type of localization of a dark, but not completely black, colour component is found as a breed feature in the Siamese cat (Fig. 12). Here, the patches are already spread out and a "mask" develops on the head. The related Birman cat (Figure 6) follows this example. Incidentally, I did not find any tendency towards the pointed colour in the the black markings of the domestic cat, but mostly saw isolated black spots in asymmetrical positions. — I have already described the particular way black is connected to the marble pattern by widening the black markings.
There are two distinct types of white with different pathological significance: leucism and albino. Leucistic white consists of unpigmented hair on pigmented skin. Albinism consists of unpigmented hair on unpigmented skin. We see both in the domestic cat, often on one animal, including with white markings.
By "acroleucism" we mean, as the word implies, the first stage of white markings, in which the white is confined to small markings at the tips of the points, in the form of symmetrical white "marks". Due to the nature of this condition, not only in domestic cats, that markings can be expected to appear appear on the ventral midline at the throat, chest, or belly, and bilaterally symmetrical markings in the armpits and thighs. In addition to this "acroleucism," the domestic cat also exhibits "acroalbinism," and these can also occur in combination as one marking proves to be leucistic, but another to the same animal is found to be albinotic [See Note]. For example, a wild-coloured "smoke tiger tabby" whose skin is grey and where the whitish base of the hair is the same colour as the skin of the white throat, but on the other hand there is unpigmented light pink skin under the white belly patch.
[Note: Physiologically, "acroleucism" and "acroalbinism" can not be regarded as counterparts to acromelanism, because the experimentally substantiated interpretation of the latter does not apply to the other two. The onset of blackening at the points is related to their lower body temperature. I touch here on recent experimental results of this kind in connection with the Siamese cat, since this breed does not belong to the truly white ones.]
Fig. 1 g (white markings "Type 8") in KÜHN and KÖNING (1928) gives a good example of the symmetrical arrangement of several such markings. (This work does not distinguish between the two varieties of white). The opposite type (No. 1) shown there, i.e. the most widespread white markings, is characterized by remnants of the counterstain (black, wild colour, "red") on the tail. Such residues can also occur on various other parts of the body, leaving the tail and other points vacant.
A special case of white marking is found in the aforementioned "smoke cat", where the basal part of the hair is white (to greyish-white) and the terminal part is deep smoke brown, which determines the surface colouring. The same distribution of colour occurs, as already mentioned under brown tigers and marble (hence "smoke tiger, smoked marble"), but they usually lack the right degree of smoke, which means, as mentioned previously, they signify the preliminary stage to the smoke cat.
Leucism, especially "Acroleucism" (in the broader sense) occurs as a breed feature in domestic animals (ESSKÜCHEN 1. c.). In this role we find white markings already exist in the wild population (in relation to which we mention the characteristic of a "type" or a "variant"). The white to yellowish throat (or chest) and belly marking in the F. silvestris group have been considered a species feature distinguishing it from the ocreata group whose representatives, as far as we know, lack such markings. However, some F. silvestris, in local populations and in individuals, lack even a single marking, the belly spot (see Schwartz 1929 a). I will deal with these cases and with the propagation of the markings in parts of this wildcat group.
The following description of my tiger-tabby Shorthair breed will explain why the racial impact of the Nordic F. Silvestris is desirable, at least in the grey individuals of this breed. Therefore, I allow in this breed the white throat (chest) and belly spot in the style of F. silvestris. A selection against these markings would also counteract that racial impact; they have always come from that race, but have been excluded.
The possibility of a tendency to spread this white, which is disagreeable from the standpoint of typing, and possibly also of breed purity, is accepted. The fulfilment of this tendency can be countered by breeding principles.
The presence of the different levels of white marking does not yet prove the propagation as a process. A general tendency for this is especially adopted for albinotic white: "The coloured areas get smaller. One by one they disappear. Amost the whole body is albinotic," it can be stated that “there are places with a livelier metabolism,” where the darker colour remains (ESSKÜCHEN with ADAMETZ). To what extent this last-mentioned behaviour also applies to the domestic cat remains to be examined. In any case, I have found the experience of breeders on the spread of piebald white confirmed, among other things, by observation of my own brown tiger, which originally had only a breast and belly spot, which developed white in the armpits and inner thighs at the age of about 5 years [See Note]. A particularly vulnerable spot in patterned cats in this regard is the chin, which results in one of the most common colour defects that increase during the individual‘s life. Experiences with the tendency of white to spread were probably mainly aesthetic considerations decisive for the French principle of disqualifying all kinds of white marked cats with a predominance of white, in both long- and short-hair (under the nickname "Calves").
[Note: With well-kept cats this age falls into the "best years; ". The Shorthair cat reaches a relatively high age. I have known ones aged 19, 18 and 16 years old. ]
The first mentioned path of propagation corresponds to the findings of OGNEV has already made on wild cats (1. c.). From his report on F. silvestris caucasica he says: "white spots are not rare on the neck and on the inner thighs." On the other hand, this makes it evident that the white can also be absent from this geotype of the F. silvestris group, and that there is already significant variability in the wild state. The series of Caucasian skins I examined in the Berlin Zoological Museum, which have a throat patch but no belly patch, also probably belong to the same type. Perfectly unpatched skins are also found in European wild cats (see SCHWANGART 1929 a). Patches, as already mentioned, also range from white to ground-colour in this group, as shown here, for example, in Fig. 4, where the throat marking is pale yellow.
When I admitted my reason for allowing white throat (chest) and stomach markings from the wild grey form and its close relatives into my "Shorthaired Tiger-Tabby" (which must remain so, for aforementioned reasons), I assumed that this wild white would not share the spreading tendency of domesticated white markings. Even though OGNEV's results show a great degree of variation already exists in the wild species, there remains the impression that despite the multiplication of the markings, the initial white areas do not expand. In any case, the increase in domestically acquired white, which apparently first appears in the smale places, must be expected in the domestic animal population.
These considerations lead back to the distinction between the behaviour of leucistic white and the albinotic white. The previously conclusion about the tendency of domestic-animal white to spread in general refers to albinotic white. However, the necessary evidence is not yet available in the wild species where we always consider beforehand that the white is leucistic. But initial signs of albinism are also found in wild animals. The zoological material of F. silvestris should be studied accordingly [See Note].
[Note: On this occasion I may refer to the interesting case of hereditary partial albinism in the lion, which M. M. SCHNEIDER recently published in 1931).]
Breeding practice arising from this situation poses the question of whether or not to permit white throat (chest) and abdominal marks in domestic cat breeds, and for which breeds, should they also be allowed a limited degree of underarm and inner thigh white, and in order to fully reconcile them with the wild group cross-breeding is desired. The obvious requirement of completely excluding albinotic white markings cannot be undertaken at the present stage of cat breeding. In the case of pure white cats, many exhibition judges prefer, or require, light skin.
With regard to pathology, albinism is generally considered degenerative. ESSKÜCHEN has gathered a lot of evidence from the different types of pets, including albinotic piebald occurrences and "pseudo" albinos, namely those where albinotic hair and skin quality does not accompany the eye colour. He provides many symptoms of degeneration that accompany albinism: lack of resistance to the sun and the weather or against food containing a slight poisonous or irritant, poor healing of skin during skin diseases, infertility, discarding, rejection of, or decrepit offspring. In cats he lists deafness in blue-eyed whites and that albinotic white iss a lethal factor.
The overall assessment of domesticated leucism is inconsistent: "Under certain circumstances" it seems "pathological", but on the other hand there are also cases of particularly strong constitution in such animals, for example white horses that are dark-skinned are the most likely to resist tanning along with grey horses.
As already mentioned, the characteristics of full-scale albinos include not only a completely pigment-free (white, pink) skin, but also eyes with a red-looking iris and an equally bright pupil. This scenario is exceptionally rare in the domestic cat. H. C. BROOKE (1930) describes such an animal, from the Paris exhibition of the "Societe Centrale Feline de France" in that year. As far as I know, this is the only documented case in the domestic cat. Up till then it had been supposed that cats did not exhibit pure albinism. The eyes, which I have just described, alone should be termed "non-pigmented," a term which is often applied to the blue eyes seen in suckling kittens and in particular breeds. I know of pigment-poor tints: a blue iris with a normal dark pupil where the pupil shines red, a case of a pale lemon-yellow iris with an intensely red shining pupil [translator’s note: "the elusive pink-eyed dilution"], gradations from greenish-blue, light-blue, milky-blue through to "Cornflower" blue. All these phenomena are due to the restriction of pigment, its nuances in the particular way it is distributed in the eyes - this is especially true of the levels of blue – and also the structure of pigment-free parts, which precede a pigmented layer. I will not discuss the details here. As far as the cat's eye is concerned, I refer the reader to the works of A. RASELLI (1923), HANS RlCHTEE (1928), ERICH Murr (1927 and 1931).
Little is known about the general constitutional weaknesses among white cats in contrast to the frequent complaints about the delicacy and farilness of the young animals and the like in weakly patterned to single-coloured flavistic animals. The data taken from ESSKÜCHEN (1 c.) By ADAMETZ (1926) on a lethal factor being related to a white factor in the domestic cat is based on observations made by other authors. It reads: "In cats, CREW suspects such a factor in a certain form of the white colour variety. He concludes this from the small litter sizes in white cats. Evidence supporting this assertion was provided by JONS (1922), who proved that pregnant white cats contained a larger number of dead embryos than pregnant coloured cats." According to him, there is a lethal factor coupled with white hair colour which destroys life even in heterozygotes. "In the cat, as in the mouse and the horse" and many others "there are lethal factors coupled with certain forms of white hair". Since the work of the authors mentioned by ADAMETZ is unknown to me, I can not judge what they mean by "certain forms of white hair." The inidicates that they do not simply mean albinism.
In contrast to signs of general constitutional weakness, a well known defect specific to white cats is deafness – a high percentage of blue-eyed-white cats are deaf. PRZIBRAM (1908) has found that in the case of white cats with differently coloured eyes, there is deafness only on the side that has the blue eye. (Of course, as well as blue-eyed whites that have hearing on both sides there are odd-eyed white cats with good hearing on both sides). Blye-eyed whites show all the stages between good hearing to total deafness. Even odd-eyed whites have a higher incidence of deaf individuals than coloured cats.
In my sphere of activity I am in favour of the orange-eyed whites rather than the current breeding specialization for blue-eyedness where the associated deafness, a defect particularly bad for cats, threatens to become a breed characteristic. In the United States, both varieties are held in the same estimation. In France, at the Paris International Cat Show in 1931, at my request, a "class" was introduced for orange-eyed and amber-eyed whites, breeds it did not previously have.
The connection of blue-eyed-white with deafness is well-known from the White Longhair. White Shorthairs are not much bred. Unanimously, however, the English standards (MORTON "Domestic and Fancy cats" pg. 13, SIMPSON pg. 101) also demand blue eyes in white shorthaired cats. To solve the entire interconnection, the contrasts of Longhairs and Shorthairs and albino-leucistic whites should be used. Consideration should be given to the blue hues, in which clear and azure blues are preferred, and greenish tints should be rejected.
With regard to the mentioned red eye-shine in the pupils of blue-eyed whites, this may be strong or weak or may even be absent. One breeder has the impression that the intensity of this eye-shine is linked to the tendency to deafness, but others deny this.
To conclude our knowledge of white in the domestic cat, the crossbreeding experiments of KÜHN and KRÖNING (1928) serve as good material. The complementary colours were "red, black and agouti" (wild colour), the results probably also apply refer to blue. Overall, the type of a "monohybrid crossing [first cross] showed intermediate features in the bastards [hybrid offspring]." The white markings are based "mainly on a pair of genes", but the degree of piebald also appears to be influenced by something else. We still await the solution to the question of whether that additional factor is a third allele to the two previously determined or is a "modification factor" belonging to another pair of alleles. In line with previous researchers (WRIGHT 1917, CASTLE 1925), the authors also show a spotting phenomenon within the "dominant white" of domestic cats and cite it as "spotting-like characters": "Loose groups of dark hair in certain places" and the presence of blue as well as coloured eyes, together with both colours on one animal or "partial depigmentation of an eye" (strictly speaking = poor pigmentation, see above on albinotic eye colouration), the latter statement being new to me. Experiments are also underway to test for this presumed "even lower level of pigment dispersal." As mentioned, no distinction was made between albinotic and leucistic whites.
In the preceding remarks regarding the occurrence of white with other colours and its proliferation over them, I emphasized the flat pattern. On the other hand, these facts do not contradict the modes of the (even piebald) appearance of isolated elements, of single hairs and groups of hair, and the spread of white over other colours as well as the fading of those other colours into the background in favour of the white colour. You will understand their presentation better in the descriptions of each other colour types that will be into considered.
The strongly tinted flavism belongs to the "dilute" or "rarefied" colours, which are characterized by a reduced level or density of pigment compared to the full colours. It is related to white, especially to albinotic white, and is also considered a precursor of it.
According to ESSKÜCHEN, the constitutional affinity between flavism and albinism manifests itself in a weakening of the skin accompanied by strengthening and enlargement of the hair. Pale yellow hair is thicker then dark brown hair. The increase in thickness in albino hair is even stronger (according to WIEDMERS). ESSKÜCHEN sees these changes as signs of a decrease in the constitutional strength already occurring in Flavism, but does not mean complete degeneration. To my knowledge, the relationship between hair colour, skin colour and hair strength in the domestic cat has not been tested.
The Siamese bears witness to relations between albinism and flavism. Siamese dilution is associated with the low-pigmented eye (blue type with deep ruby glimmer of the pupil), and newborn Siamese are white; the colour change to cream, along with the characteristic smoky masking (Fig. 12), occurs during the first few months. The process of change develops in such a way that the flavism emerges from an almost albinotic state. Many adult Siamese show traces of a marking pattern, and this will have been present before any domesticated colouring. It is probably permissible to infer a tiger pattern.
Our native short-haired and long-haired cats in "red", deep yellow, Isabelline, and cream show a tiger or marble pattern in most cases. At least only in those rare transitional stages, where the entire hair is paled to near white, is the absence of any striping the rule.
The albino relationship is revealed in the following case which we considered when enumerating pigmented eye types. The animal, a foundling, was very pale, dull cream colour, against which faded tiger stripes stood out on the torso. The iris is a pale lemon colour, the pupil has an intense red shimmer. Against my interpretation of this case as a Flavistic-Albinistic mutant with the peculiarity of still consistently recognizable striping, one could consider the possibility of a Tabby-Siamese hybrid such as TJEBBES (1924) has bred from a crossing of a grey tiger-tabby and a Siamese. However, our specimen also shows special signs of degeneration: her eyes are hypersensitive to light, and so far she has been in heat only at abnormally long intervals, and in those cases so inadequately that the hoped-for breeding attempts (mating with Siamese, red tabby, and others) could not be carried out [see note]. But it is mentally normal and is especially sociable and lively, but not irritable.
[Note: A small degree of hypersensitivity to light is also exhibited by many Siamese. The "squinting", pink-eyed condition is often even considered a "race characteristic", and is not to be confused with the widespread strabismus fault spread by selective breeding. On the other hand, I have not noticed this tendency with blue-eyed whites, and the Siamese also lacks the tendency to be deaf.]
Regarding constitutional weaknesses in "self colours" and in the isabella and cream colours showing rudimentary patterns, these were usually noted: rejection of young, poor rearing, decrepitude of the young, and susceptibility to distemper. Whether the statement made to me is correct, that England succeeded in gaining breeds free of such weaknesses, I cannot judge.
Most conspicuous among the Flavistic cats are the Siamese, with their general and also special constitutional defects. As already mentioned (SCHWANGART and GRAU), I associate the gross exaggeration of strabismus [cross-eyes] and the various degenerative tail deformities in this breed with inefficient treatment together with poor selective breeding. These animals are considered by many owners to be "tropical" and exposed to harmful treatments, such as being shut away from fresh air and sunling, and overheating, although good strains have been bred for many years in Europe. In additiion we have the conscious selection for those defects. Apart from all this, however, they have an inherent tendency to the defects mentioned, such as a general constitutional weakness, which often causes difficulties in their breeding. Well-bred adult animals, on the other hand, are full of vitality.
Experimentally, little is known about the inheritance of our homegrown "red" dilution as such. BAMBER (1 c.) establishes its character as a "dilute", and subsequently puts these nuances (see her "Note" pg. 14) together with the deeper hue that contrasts with black and blue in her analysis of "tortoiseshells". The group is called "yellow" and includes "orange" (our "red") and "cream" (probably including our "isabella"). In the tables reproduced by BAMBER after WHITING, a "cream" occurs only once, but again it represents the group and is not compared with a full colour. In view of the aforementioned rarity of self-colour deep "reds" it would probably be useful to try crossing the almost "full colour" red tabbies. These are also certainly called "orange" by WHITING.
The special case of "Siamese dilution" enjoyed more attention in the inheritance experiments. Despite the breed’s partial albinism, this appeared as an allele opposite white, where white dominated in the same way that "pure white" appears to "dominates" over all other colours (and patterns). In addition to crossing with white, the Siamese colour has been bred to the grey tiger pattern, yielding the "completely new type of striped Siamese" which I have already mentioned (TJEBBES 1924). Racial breeding would be detrimental to the appearance of these mongrels, incidentally they do not take after themselves [breed true]. The Siamese was not crossed with "deep red." On the other hand, it was used for infestigating the inheritance of eye colours. Mrs. BAMBER wrote about this point: "In truth we have no single fact concerning the inheritance of eye-colour that we might even call reasonably assured, and in almost the majority of cases our ignorance is perfect."
The other "diluted" colour is the blue ("Maltese Dilution", as a dilution of black). Like the solid colour (see above F. silvestris daemon SATUNIN), so also this dilute colour also appears in wild forebears; E. LÖNNBERG (1922) found it in the coat of a F. ocreata from Erythrea. He also mentioned the case of a blue tiger (Uncia tigris).
I have already reported on the tendency of the Blue Shorthair to have a slender form (a tendency in which it is second to the black), and the opposite deviation in the case of the Blue Longhair which tends towards the Persian type. From such signs of constitutional "break-up" and "disturbance," we might also wish to infer that this colour produces degenerative tendencies. In contrast, I have so far found no evidence of inferiority, considering the pronounced popularity of both Blue Shorthairs and Blue Longhairs among breeders in the balance. Also the English studbooks, which are worth consulting in the case of such actualities, contain, as far as I have seen, no information about particular difficulties in breeding and keeping of blues. But perhaps this is related to the requirement for "blue skin", which has long been established for the Longhair by default and which applies, at least traditionally, for Blue Shorthairs.
On the other hand, there are difficulties in keeping this colour pure, and even here the blue appears "unstable". A distinction is made between two basic shades, the "steel blue" that is closer to black and the "silver" blue (pale blue); both of these are permitted by breeders. The second colour, as good as it is in the Blue Longhair, in the still early days of German short-hair breeding, is complained about among breeders because of its tendency to whitish fading and the appearance and increase of initially scattered groups of white hairs. The steel blue also has a tendency to approach the silver blue colour.
The silver blue follows the previously mentioned actual silver colour, which is very rarely found as a solid colour, but is most often associated with markings and already associated with wild colouring, but on the other hand, in certain cases (see above), it is found in association with white.
While blue tiger-tabby and blue marble are not rare combinations, I have so far found the basic colour to be pale blue and the pattern to be a deeper blue to black colour. I have not yet seen either of those last blue shades as solid colours directly against white or against black, while, as previously mentioned, transitions from diluted "red" to white are even traceable as occurrences in an individual. On the red scale there is a not a continuation between deep orange ("red") and black, but there is a foxy red-brown in the place of black. Sprinkled white areas of hair are common with blue, as well as a white throat (chest) and belly, but extensive blue white spotting is less common.
Breeding practice provides as ready a corrective in the crossing with black against the fading of the blue. According to WHITING (1918 and 1919), blue behaves "recessively to full colour", this was based on a "uniform inheritance factor" (BAMBER). A solution would be found to the apparent contradiction between a breeding practice that is rich in experience and the scientific cross-breeding results if the result of the practice was not a union of the two colours, but a restorative effect of the crossing process as such, in the appearance of increased pigmentation, such as ESSKÜCHEN (see above) mentioned (von Lämmern).
Here I refer back to my previous assumption of the role of blue in the establishment of the "Annamese breed" (which I have not seen) which were the colour of Blue Shorthairs with the shape and mask pattern of the Siamese.
In my account of results on the individual hair colours, I have dealt particularly closely with crossing experiments. I will now proceed with an overview of attempts with crossing of features from various colour groups. What has been achieved so far in this field is still unsatisfactory, and to my knowledge, renewed investigation is under way, following the aforementioned experiments with white-markings. For details I refer to BAMBER’s dissertation (1 c.) where, for example, 30 out of the 86 printed pages deal specifically with tortoiseshell ("tortoiseshell") spotting, which, as I said, is not considered "race-forming" in accordance with my theme.
First of all, with regard to the simple colours and the two marking patterns, the reference to the prevailing (according to the numerical ratio of usually "imperfect") dominance ratios: white dominant over all other colours and patterns, patterned is dominant to black, tiger-tabby is dominant to marble-tabby, black is dominant to blue. Regarding the behavior of black to "yellow" (which was usually patterned in degrees) the tortoiseshell marking comes into play, in which blue or a marking type can represent black. As a rule of producing tortoiseshells, black-and-yellow crossbreeding (or blue or grey-patterned with yellow) to black (or blue, grey-patterned) and yellow tomcats, with a tortoiseshell female will produce them. In my experience, the pattern of red or yellow shades is easily lost in the tortoiseshell pattern (in the broader sense), with the result of producing a uniform tint, whereas it is preserved in grey tones.
In multiple markings I have seen black, white, flavistic tones right up to "red", blue, and all sorts of tiger and marble patterns side by side. The distribution of these colours and tones may be solid or small amounts may be mixed together, or they may form scattered patches, especially with white. These distribution types can occur next to each other on a single animal. As a rule, however, specimens without white (tortoiseshells) have a small mixed patches of colour, looking rather unkempt in the Shorthair, while those with the addition of white (the so-called "Spanish" type) having an arrangement of larger patches with a tendency to smoothness and glossiness in Shorthairs, which is desirable in breeding.
The standards recognize only animals with black and reddish-yellow tones, as "pure tortoiseshell," and recognise those with reddish yellow tones, black and white as "Spanish." They require distinct patches of colour in both types which, as I have said, is a rare exception in the "pure tortoiseshell." According to SIMPSON, the "tortoiseshell" pattern must be "spread out, like a turtle shell," from which this type gets its name.
I have already mentioned in the section on flavism that Mrs. BAMBER, in common with her sources, treats all gradations of the red-yellow group right down to the pale dilutions as one factor. Accordingly, this lacks a proper definition of "tortoiseshell", whether this type is based on three colours, as indicated by cat breed books - where two of the colours are represented by different shades of yellow-red - or whether two colours prevail or suffice with just a nuance of the "yellow" factor.
The studbooks treat the tortoiseshell as "tricolour" and leave out any indication as to whether bicolour piebalds of this variety ever occur. “Domestic and fancy cats” (1921) says of the Shorthair: “tortoiseshell is black, yellow and red”, and of the Longhair : “black, orange and yellow”, the difference in colour terminology is due to loose language. SIMPSON speaks of "black, orange and yellow". (Blue and patterned versions are undoubtedly not mentioned because they are not allowed in breeding).
The black and yellow tortoiseshell I saw usually had at least two reddish yellows hues present, and sometimes more. With the predominant character of a colour mixture, it is difficult to find a consistently sharp distinction between the tones. The "Spanish" piebald with its wider-spread colours often has enough uniformity for us to determine the yellow-red components. According to the genotype of the real tortoiseshell, taught by experiments, they should only occur with a red-yellow shade, which means these animals have two colours. The colours resembles those in the Siamese pattern, which is not inherited in a gender-linked fashion, and which has a different genotype.
Many already consider it necessary for breed-knowledge to make a connection between experimental heredity research aimed at determining and clarifying the rules of inheritance, and the investigation of the development and characteristics of the research objects. In his results on hair colours, W. JANKOWSKY (1 c.) rightly sees progress towards a better understanding by deriving a postulation of its "genetics" that will have to be used following successful proof of a "hormonal basis, of oxygen and peroxidase as real factors of pigment formation, instead of the previous speculative factors."
In my remarks on colours as well as those on patterns, I have cited certain kinds of transformation, and I have always tried to respect the difference in the testimonies provided, whether they manifest the change in an actionable way - individually, over the course of generations - or only suggest it by being an intermediate state. Such events were detected directly several times.
From the point of view of research into the causes of such processes, which I have already dealt with, the majority of our cat population occupies a special position as objects of investigation because a number of domestic actions which otherwise accompany pet ownership have not yet happened to it. This exceptional position is even more favourable because there are again groups of the same pet, with similar colours and markingss, which were exposed to domesticative influences, the ancient Persian and Siamese breeds. The current keeping and breeding of cats probably gives us the best possible opportunity to see those influences at work.
Here we come to the topic of inbreeding. ESSKÜCHEN (1 c.) makes this, as I have already pointed out, partly responsible for the fading of colours, but thinks its damaging influence is limited to the group of individuals, as normal dark animals reappear on crossing or introducing new blood. To solve this question, it would help to study bleaching in uncultivated Blue Shorthairs compared to bleachining in strains of blue Persian subjected to decades of close inbreeding.
On the question of the influence of lifestyle on colouring, the neglected cats provide their contribution, showing how hair quality, style and lustre suffer severely and how its colour tends to fade. They also show they can recover their original appearance through gentle handling, correct husbandry and care. Even more than the often inadequate and usually unwholesome food, since domestic cats have a passion for catching rats and mice for nourishment, other conditions of life that are unfavourable to the pet will take effect, not least the psychological insults that have a shocking and serious ill-effect on the cat’s constitution, of which I have many examples. [See note].
[Note: Outcast domestic cats focus on pursuing their goal of regaining a home in the human community. There is another possible tolerable state to which some cats can adapt psychologically. This results in the complete abandonment of man and corresponds to the original wild state. The widespread assumption that the wild colour will then be recovered from pet-coloured specimens has not be proven and this leads us to the questions touched upon here.]
ESSKÜCHEN establishes an example of colour-fading due to external influence i.e. the "seasonal colouring" of grazing animals, which is produced by intensive sun-exposure. It occurs to the most affected parts of the body. This case concludes with the results of experiments with the category of "incomplete albinism," which the case of the Siamese cat belongs to. In conclusion, E. FEIGE has just reported (1931) on new experiments by N. K. and V. N. ILJIN on the Siamese cat in parallel with those of the Russian rabbit and other species. On the Siamese cat the colouring of adult animals can be chnged by temperatures – darkening in cold weather, brightening in warmth - and also by local treatment in individual places. In other species X-ray radiation achieves similar effects. Similar effects in domestic cats are caused by unintentional domesticated "experiments", except that in these cases different environmental factors usually act at the same time. I have already mentioned "rusty" black cats. A brown wild coloured Shorthair tabby of mine, with otherwise consistently harmonious colour, shows the same rustiness in places, which greatly impairs his appearance, after predominantly staying indoors during the winter (he otherwise usually stays on the sunny balconies) and at times of increased "spraying;" the same thing also happened to him during a serious illness. This colour change, therefore, also corresponds to a weakening of the constitution. Also noteworthy in this respect are the fluctuations in colour in pregnant and nursing cats, especially in the case of large litters, phenomena which need to be studied more closely.
In the section on white colouration in the domestic cat, everything indicated a lack of a sufficient distinction between leucism and albinism. This goes beyond the importance of studying the skin colour next to hair colour. So I mention here a worthwhile counterpart to the questionable desire of breeders to have pink skin in the whites and "blue" skin in the blues. Using the example of the roan in horses, which is considered pathological, we notice a change between pigmented and pigment-free areas in dark-born animals, which gradually lose pigment and become white. In the "pinto pattern" (leukoderma) of the horses we find coloured areas of hair on pigment-free skin. In the domestic cat, different levels of skin pigmentation are found, and each one of them occurs in conjunction with various hair colours.
For the time being only random samples are available. I have chosen one found in the population of a cat home and added two cases from my own animals. Here there are already a wealth of possibilities, and there is still much more to be discovered. Here the terms "Tiger" and "Marble" refer purely to the marking pattern and say nothing about the conforation of the specimens. It just so happened that no pure white animal was available. Absence of the valued blue animals is norm in cat shelters. All animals mentioned here were shorthaired.

1. Hermaphrodite: hair colour black. Skin colour is deep iron-grey throughout.
2. Female: hair colour black. Skin colour deep gray.
3. Female: same as the previous one.
4. Female: hair colour black, with white breast-star and belly spot.
Skin colour under the black hair was white, and under the breast patch and belly was pink.
5. Female: (juvenile, Slender Breed), hair colour black, with trace of white throat and a white belly spot.
Skin colour: Whole of the back was deep blue-grey, slightly lighter under the black hair of the abdomen, bright reddish-purple under the throat-spot, pink under the belly-spot. The borders of the colours under the two white markings adjacent to to the dark skin coincide sharply with the borders of the hair colouring.
6. Female: (Juvenile), Black and white piebald.
Skin colour under the black hair was dark purple throughout, the skin under the white was violet-pink.
7. Female: black and white piebald. Deep grey skin colour under the black, very light dull grey skin colour under the white.
8. Female: Black and white piebald, predominantly white. Skin colour under the black of the neck and under the dark dorsal patch was blue, and generally overlapped the white-haired region, it was pale violet under the white hair of the back, the skin of the chest and front part of the abdomen was violet-pink, that of the thigh, inner thigh and the area between the thighs was bright pink.
9. Female: Hair colour white with wild-colour tiger-tabby patches. Skin colour: Pink under all white haired areas, deep grey under the wild-colour tiger-striped hair.
10. Hermaphrodite: red tiger-tabby and white patched. Mainly white hair with a very slight yellowish tint. Skin colour: Dull grey under the tabby, dark purple under the whitish areas.
11. Female: three-colour piebald: Partly pure white, partly slightly yellowish white, in transitional areas more strikingly yellow, grey patches. Without black. [Translator’s note: blue-cream and white, with cream areas showing slight tabby markings.] Skin colour an even deep purple throughout.
12. Female: tortoiseshell. Hair colours mixed densely together with no larger patches of colour. Skin colour: Consistently light violet even under the black hair.
13. Female: tortoiseshell, with small white breast and belly markings. Skin colour: Bluish-white under the black hair, deep purple under the breast marking! Pink under the belly spot marking.
14. Male: Red tiger-tabby with white chin. Skin colour under the general colouring was gray, that under the white chin marking was pinkish-purple.
15. Male: Isabelline (pattern very weakly indicated, probably marble-tabby). Skin colour white on the head, pale bluish-white behind the head, white on the back, pale-bluish-white on the abdomen and thighs and white between the thighs.
16. Female: wild-colour tiger-tabby (full grey-yellow wild colour, interspersed by sole-stripes, i.e. a state between that applicable to the silvestris and the ocreata group.) Also the physique and head of this animal are "correct"). Skin colour! The neck and back are lead-coloured, the back is light grey, the middle abdomen has a pink tinge, the back is pink, the inner surface of the thighs is white. A white belly patch in the hair, however, does not exist on the skin. Under a narrow white hair border, the lower lip is contrasting pink.
17. Hermaphrodite: Smoke tiger-tabby (white basal part of the hair, deep brown at the tip, as deep as, but not brighter than wild colour). Skin colour: grey throughout.
18. Male: Brown wild-colour tiger-tabby (with light grey basal part of the hair), white throat and white belly spot. Skin colour: Under the main hair colour it was grey throughout, under the throat spot it was a slightly lighter violet-grey, under the belly spot it is a continuous contrasting pink.
19. Female: Wild-colour marble tabby. Skin colour: Light blue under the black pattern, light violet-grey under the background colour of the back, pink on the breast, greyish blue on the neck, pink on the abdomen (without white hair), pinkish purple on the inner surface of the thighs, pink under the pale yellow chin.
20. Female: Wild-colour marble tabby. Skin colour: white on the back under the black and grey hair, bluish-pink discoloured white on the middle of the abdomen, bluish white under the lighter colour hair of the throat, pink at the back of the belly under wild-colour hair, violet on the inner surface of the thighs.
21. Male: Silver marble tabby of consistent uniform background colour with strongly contrasting pattern, also by the way typical "marble-tabby head", a beautiful animal.
Skin colour: Under the black pattern as well as under the silver background colour is uniformly blue, changing slightly in intensity, with the blue on the back gaining in depth and that on the inner surface of the thighs and between the thighs having the lightest tint. The skin colour of this exemplary animal is also harmonious, unlike the equally good wild coloured tiger No. 16.
22. Male: Grey marble tabby.
Skin colour: The skin was white on the back under both the pattern colour and background colour fur, on the belly it was slightly bluish.
On the basis of this sample, it can probably be said: In skin colour there are transitional nuances of pink and white over violet, grey, blues of varying intensity through to deep iron grey and steel blue. This scall also includes transitional tones between the characters addressed as "albinotic" and "leucistic.". A yellowish or brownish skin tone, which may correspond to "red" and flavistic hair, was not found in these samples. The nature of the coincidence of hair and skin colouring is often paradoxical. Not only are there dark tones under white hair (leucism), but also white skin under both black and light tones, and indeed albino pink skin under wild colour hair and wild pattern coats. The areas of skin colour, which often change several times on the same animal, can be marked by sharp bounderaries matching the hair colour boundaries, or can spread over the hair colour boundaries. In some of cases, there is uniform skin colour beneath wildly inconsistent hair colour, and in other cases we see the reverse of this.
Special consideration is needed for practical breeding of those cases in which, in which appropriate skin colour goes together with markedly good quality colour and markings (as in example No. 21). In the grey shades in patterned cats, marble-tabbies as well as tiger-striped, including the basic silver and blue colours (for which the same requirement already applies), we are almost ready to demand that their skin colour must be consistently dark and as uniform as possible. This require can probably also be extended to the two "Wildcat white marks" permitted in wild-coloured and related tiger-tabbies, even if it should be pointed out that even in F. Silvestris, in addition to grey-toned skin, the areas under these white markings have pink skin. It seems desirable to apply the same requirements (dark skin), among others, to the red tiger-tabby and the red marble-tabby, in the expectation that this property would counteract the fading of the background colour and the weakening of the markings.
Undoubtedly, problems of laws remain here which are scientifically noteworthy. For the time being there is no indication of age-related or developmental tendencies (as is seen in the progressive roaning in the horses, which has not been observed in the domestic cat, at least not to the same extent). We consider it certain that skin colour, in combination with hair colour, has inherently constitutional importance, and not merely in differentiating between leucism and albinism. Among other things, the difference between pink and white skin needs explanation.
Since the majority of such shelter cats are malnourished, often severely stressed, having arrived as strays and foundlings, we can expect symptoms of such conditions to be reflected in the skin colour. This may result in cases of abnormal bleaching of the skin (such as anaemia and loss of pigment), so that light grey is bleached to pink or even white, and blue is bleached to violet. The main thing is that these circumstances do not permanently change the result.
Fig. 1 in the work SCHWANGART and GRAU (1931) showed a hairless cat with purely albino skin and noticeable symptoms of deterioration. (Its eyes were not albinotic). In this year's Paris International Show, I saw another specimen with vividly patched skin: pink and different tones of violet to blue, the areas clearly defined with sharp boundaries ("tile-like"), just as I saw in the furred specimens listed above. In contrast to the appearance of that albino-skinned specimen, this second animal had no external signs of deterioration in posture, shape or skin condition, except that its non-albinotic-looking eyes were hypersensitive to light. The dentition should be examined. (On the whole, this hairlessness is degenerative and is not desirable in breeding). As mentioned in the cited work, it is likely that the two animals were closely related.
A parallel study of thoroughbred Persians is desirable to determine the condition of the skin colour after completion of selection for hair colour.
In the domestic cat, as well as in the dog, the skin markings extend to the mucous membrane of the oral cavity and the pads of the feet, and the colouring of the outer skin may be different to the hair colour.
A phenomenon that has never been investigated in the domestic cat, as far as I know, is the variegated colour of its nose leather. Ernst Schwarz (1930) counts the "very red nose" as one of the characteristics of "true wildcats" of the F. silvestris type, in contrast to domestic cats; I had the opportunity to see this in one of the two wild (Siebenburg) wildcats. Different degrees of red are also found in domestic cats, and Schwarz’s remark gave me reason to take a number of shelter cats for a closer examination of this feature.
Findings:
Colouring pure white (female), nose leather pink;
Colour white with grey ringed tail (female), nose leather pink;
Colour white with black nose bridge (female), nose leather black with pink middle line;
Black and white piebald (female), nose leather pink;
Black colouring with a little white, but white bridge of the nose (female), nose leathe pink;
Black and white piebald with black spot on the nose (female), pink, one-sided blackened;
Black and white bicolour, white on the nose (female), nose leather pink;
Colouring pure black (female), nose leather black;
Colouring as above (male), nose leather black;
Colouring as above (female), nose leather black;
Colouring as above (female), nose leather black;
Tiger-tabby with a lot of white, white on the nose (hermaphrodite), nose leather pink;
Tiger-tabby with white face (female), nose leather pink;
Tiger-tabby with a lot of white, face white (female), nose leather pink;
Grey tiger-tabby, with whitish lip (female), nose leather pink;
Colouring as above (female), nose leather pinkish tint;
Colouring as above (female), nasal pinkish with black shading;
Silver tiger (female), nose leather rusty red, black interspersed;
Silver marble-tabby, a little pale (female), nose leather rust-to brick red;
Silver marble-tabby, reddish (male), nose-leather dull reddish, black smoked;
Grey marble-tabby with white lip (female), nose leather translucent light red;
Wild-colour tiger-tabby with slightly white chin (female), nose leather fiery brick red;
Smoky tiger-tabby (hermaphrodite), white basal part of the hair, grey skin, dark face, nose leather blue;
Brown tiger-tabby (male) with wild colour meeting brighter basal part of the hair, white chest and belly spot, pink skin under this, nose leather black;
Pure wild-colour tiger-tabby (male), nose leather brick red;
Colouring as above (male), nose leather bright brick red;
Colouring as above (female), nose leather dull brick red;
Colouring "Spanish" with grey (female), nose leather pink;
Colouring as above (female), nose leather pink;
Colouration "Spanish" with little grey (female), alabaster-yellow nose, as if translucent;
Colouring "Spanish" with a lot of white in it (male), nose leather pink, a black spot extends from the bridge of the nose onto the nose leather.
[Translator’s note: the preponderance of females is because people tended to get rid of females (which would have kittens) and keep males (which were easy to castrate). Spaying was risky and uncommon.]
From this we can judge: here, white hair corresponds to a pink nose, and black hair a black nose. This pink and black is strongly dependent on the colour of the hair surrounding the nose leather, in this way we also get two-coloured nose leather. It probably also corresponds to the skin colour (deep black nose leather on the smoke tiger-tabby which had a white basal part of the hair, but grey skin). Different intensities of red appear with the tiger-tabby and the marble-tabby pattern, even if they are silver tabbies. This finding supports a connection between the silver colour and wild colour. We see that the red of the nose leather in domestic cats reaches an intensity that cannot be exceeded in wild cats. These striped cats are not necessarily associated with the moderate F silvestris shapes, and the same applies to the marble-tabby pattern, which is obviously as common as the tiger pattern and is not a wild pattern. One wild-colour tiger-striped young cat with bright red nose leather was far too slim to meet my breed concept of a shorthaired tiger-tabby.
I recommend restraint against the temptation of using the crimson nose leather as one of the criteria for species definition. The question of the systematic significance of this feature among the cat’s wild ancestors is still unclear. It remains to be established whether it is present in all types which belong to the Felis silvestris group, and whether it is replaced by black nose leather on all Felis ocreata, or to which of these groups it belongs to it approximately, and also whether or not it already occurs individually in the wild material. Even if, as may be assumed, the majority of the types and individuals attributed to F. ocreata have "black noses", the crimson nose might not be considered a requirement for any class of patterned Shorthair: the short-haired marble-tabby breed does not require either the pattern or body shape which I associate with F. silvestris ancestry (as will be shown here), while for the shorthaired tiger-tabby breed where F. silvestris ancestry is desireable, we do not object to a background of ocreata-type ancestors. Demanding a "red nose" may seem more acceptable for patterned Longhairs as here we consider F. silvestris to be the main type. But this also requires the examination of a large number of cats of this type. For the time being, similar investigations are necessary for long-haired domestic cats, both patterned ones and otherwise. Using the nose leather of wild and domestic cats as an example, we can see how the subtle use of individual features makes us miss other, more conspicuous, features. The F. ornata group should be used when examining the wild species.
Pink nose in the grey-patterned domestic cats as well as in the black and blue (should it occur in these two) probably means something detrimental. In the case of the redness of the nose leathers in shelter cats, I believe that the alabaster-like, glassy tone observed in one of the cats should be regarded as a symptom of weakness due to unfavourable living conditions before it was admitted to the shelter. The colouring when it was in a household was solid red.
With regard to eye colour, so far we have only addressed the albinotic group. My guidelines (SCHWANGART 1929b) call for harmonisation of hair colour and eye colour in accordance with their tendency to occur together. It thus meets the aesthetic point of view and generally agrees with the foreign standards. The exception is the case of degenerative blue eyes in whites. Combinations are: green to yellowish eyes in the wild-colour tiger-tabby; green eyes in the slate grey; greenish when silver striped; greenish or lemon in the bleu striped; orange or amber in the brown-striped; yellow or orange in the red striped; the same combinations in the marbled tabbies as in the striped tabbies; deep golden yellow or amber in smoke cats; in white cats deep yellow (golden, orange) is better than blue; orange in black and blue cats; green in silver and related colours; deep yellow in red and isabelline cats; in bicolours it corresponds to the predominant body colour; orange to light brown in tortoiseshells; orange in "Spanish" colour cats. These rules, which are applicabe to Shorthairs, are aligned with those for longhaired cats which are more richly coloured due to selective breeding. Scientifically and practically desirable is an investigation into the relationship of eye-colour to skin-colour and to the distribution of colour, and to the influence of the colour immediate surrounding the eyes such as is shown in the case of nose leathers.
In addition to the aforementioned knowledge of colouring are the colours of the bare paw-pads and the claws. The colour of the paw-pads, which may be black or pink, is included in K. M. SCHNEIDER's "Case of Hereditary Partial Albinism in the Lion" (1930a). There is a close relationship between the colour of the naked parts and the adjacent furred parts. The claws are blackish and whitish in the domestic cat. English standards demand "white claws" in Chinchilla Persians.
VI. MY SHORTHAIR BREEDS AND THEIR SYSTEM.
Now that I have completed my descriptions of patterns and colouring in the domestic cat, I will return to its conformation. According to the English breeding principles we have alreay seen features of form as the basis for establishing breeds, and in this case the Longhair taking precedence (whereby we fall into the one-sided, exclusive cult of the "Persian") and it is a determining factor in the Siamese and its new long-haired relative, the Birman, and under the domestic Shorthairs in the so-called "Abyssinians" and the Blues, however, shortcomings and differences of opinion.
The shape of the domestic Shorthair can now be recognized as two almost opposite extremes, without the various parts of the body always needed to agree.
On the one hand we have: stocky trunk, sturdy legs, short, broad neck, which makes the head posture more level, broad forehead, short face with wide blunt snout, thick, relatively short, slightly bushy tail (fig. 20 to 22). But on the other hand: (Figs. 23 to 26) Slender body, long-legged, slim, delicate, graceful and even slightly curving neck, which carries the head high, narrow head with a strongly tapering, sharply pointed muzzle and a somewhat arched (but not protruding) forehead, the eyes typically inclined ("Mongolian", see Fig. 26), long, thin, wholly short haired tail.
The example of the "Abyssinian" and that of the Blue-haired ("British" and "Russian" Blue) has shown that it was the consideration of such differences that subjugated English breeding when it came to establising these varieties, although this consideration remains uncertain, inadequate and haphazard.
In my works (1928 b and 1929a) I tried to show that similar contrasts may have been regarded as racial characteristics in ancient Egyptian populations and that they were initiated in local forms of the wild population (F. ocreata), probably individually. Discoveries of skeletal remains confirm the ancient Egyptian depictions of the (culturally conditioned) ideal of a thin breed follows the same limitations; L. ARMBRUSTER (1921) claimed that in the case of the ancient Egyptian dogs, according to which "we may take it for granted may (excessively slender) forms as in the illustration 'desert animals' . . . are not quite true to nature." However, the existence of the opposite form, together with the intermediate forms, was long since proven by domestic cat discoveries from that time (by authors of the Berlin "Discussion of the Ancient Egyptian Domestic Cat" 1889, LORTET and GAILLARD 1903, in addition to my aforementioned brochures). In addition, this is also revealed by comparing the ancient representations (pictures, statuettes). It was recently proved by I. AHARONI (1930) that these pieces commemorated the different shapes of wildcats that coexisted in the region it inhabited, He says that "both" Felis ocreata bubastis Hempr. and Ehrb. and F. ocreata maniculata RÜPP are "spread throughout Palestine and the East Bank." He could "not set firm limits on the occurrence of both. Both occur in Egypt." The question then arises as to whether these are really species differences or individual differences.
Of the two contrasting Shorthair complexes, it is certain, on the basis of our knowledge of Egyptian wild cats, ancient Egyptian domestic cat skeletons, and the ancient depictions of cat, that the one leaning towards the a slender form seems to have been perfected by the ancient Egyptians, but not the one tending towards the stocky form. The conformation of the second type mentioned corresponds much better to the characteristics of the Nordic F. silvestris, as well as to the characteristics of the long-haired breeds, whose description has already led me to discuss the question of race and the resulting affirmation. The stocky Shorthair breed conformation, as I wish to read it, corresponds especially to the German Longhair. Compare Fig. 7 (F. silvestris) 5 (German Longhair), 20 to 22 (Shorthaired Tiger-tabby). Therefore, we have on the one hand, in the case of the naturally initiated Shorthair, the slender type, probably the result of ancient selection, and on the other hand we have the contrasting stocky type, perfected by a stroke of luck through free crossbreeding with a second wild-form. Thereafter, all those sub-characteristics of the above characteristic innately belong to those other on both sides.
Genetic studies on the inheritance of individual characteristics as well as inheritance of whole complexes are still completely absent; in the case of the domestic cat studies are restricted to preferred features: colour, to a lesser degree markings, and, as far as form is concerned, to malformations. Inheritance science and older breeding practices are in agreement. The boundaries of normal conformation features are more difficult to understand than those of other features.
With regard to the breedability of both contrasting conformation complexes, I believe that I can say from practical experience that in the case of their characteristics they are inherently inclined to go together. For this purpose, reference should be made to the relative rarity of pure-breeding domestic Shorthairs, which applies to conformation as well as to markings and colours of random-bred strains. (However, I have also seen homozygosity in complicated pattern-colour combinations in the random-bred population.) I again tentatively give the impression that in cases combining features from both confomration complexes, mutual individual characteristics (sub-characteristics) usually appear weakened, whereas the harmoniousness of the detailed characteristics becomes more pronounced.
Regarding the proportions I have this to say: The compact from is the most widespread, but there is also a large percentage of medium size cats, and quite a number of very small ones. Also, lean animals are quite large, but animals with first-class conformation are less common in these than in medium size cats. We don‘t require two two types of Shorthair when breeding for size. Stocky build, the massiveness of a good long-haired cats would be desirable. In fact there are isolated cases of stocky short-haired cats reaching these dimensions; this is proven by depictions of short-haired tiger-tabbies by SILVESTER (Berlin) and SIMSON (Vienna) which arre shown here, both of which are larger than some of the F. silvestris specimens I have seen.
With regard to the two contrasting forms, as presented by the Shorthair and how desirable they are in terms of breeding, the question of their relationship to the normal limits is relevant. While, as already said, the Persian Longhair must already be considered excessive, in the case of Shorthairs, in the natural state the stocky conformation commonly adheres to the normal limits; the determination of these limits is considered to include the conformation of the northern Felis silvestris, and the German Longhair is its special long hair counterpart. I have already considered the excessive tendency of Shorthairs, especially Blue Shorthairs, in the direction of the Persian head, to the point of becoming pug-headed, and have dealt with the question of their breed potential. In specimens of my basic stocky form I would be agreeable to the shortening or widening of the muzzle beyond "Silvestris limits" as long as it is not a blatantly excessive features.
In contrast to the stocky conformation complex, I have already discussed the extremely developed slender head in the essay SCHWANGART and GRAU (1931) as extreme conformation. Inthe same way as the dog it is clear to what extent breeds are based on the narrowing and lengthening of the head. These are worthwhile lessons to apply in the case of the domestic cat. It also needs to be studied regarding the kinking of the craniofacial axis as in the case of extreme conformation in dogs. Regarding domesticated characteristics of their dentition, the work of R. F. SCHARFF (1906/07) is a starting point. The comparative representation of skull and teeth in wild and domestic cats by JOH. BUNGARTZ (1896) cannot be considered decisive because, although it excellently offers the typology of the "European wildcat" it ignores that of the "domestic cat" the typology unconsidered. POCOCK's finding (1916) of noticeable differences in the skull and teeth within an (isolated) geotype, the Scottish F. Silvestris is significant.
I now come to the description and justification of my additional restrictions to the above two conformation types of the native Shorthair. It is possible to consider additional details about form and to group together the features of the design with the markings and colour. First of all, I would like to remind you of my fundamental remark at the beginning of this work, that the foundation of breeding lines should be governed as much by aesthetic, pure-breeding, utility-breeding, psychological and ethical motives as by evolutionary history and inheritance laws. In the combination, which leads to my listing of Shorthair breeds, aesthetic and pure-breeding considerations are considered first.
In detailing the components of conformation we must restrain ourselves in these early days of selectively breeding a native Shorthair. The breed founder himself falls prey to misleading impressions on this point, or on the general connectedness and fixability of such details, and there is the danger of getting distracted from the main features and unduly complicating a desirable basic image. I have already mentioned the backwardness of breeding technique, the various external obstacles, and the lack of desire to breed Shorthairs. Once we have set the basic type, there will be no lack of proposals for "refinement" in this breed just as in the older lines of animal breeding. The view cautiously expressed here may seem crude to future breeders.
It seems necessary to further refine form details, even at this stage when the contrast of forms is just one such general negotiation after the fact. The animal breeding code of practice is justifiably concerned about breeds based on form alone. However, loking at Figures 20 to 26 as well as at the analysis of conformation given above is sufficient to remove this objection against our case.
Additional form details I have considered so far:
Firstly, the curve of the nose bridge. I prefer a slight hook [i.e. a stop or depression] in the stocky type (Fig. 3a), but a straight nose is acceptable (the same applies to the German Longhair, see above), in the slender form there should be a slight saddle (Fig 3 b), but again a straight nose is not rejected.

Secondly, I allow both types smaller ears than the English standards allow, the Shorthair requiring "big ears" just as Longhairs require "small-ears," with the short-haired "Russian blues" already being an exception. (I agree with this regarding Longhairs, and also agree for Shorthairs except for the two types in question here. For F. silvestris, as already mentioned, "small-eared" is correct, as is "large ears" in F. ocreata, which we can be confirm to be African). The tendency, especially in tabby cats to be stocky with small ears (refer to Fig. 22) is, in my opinion, based on the impact of F. silvestris blood, the opposite tendency in the slender form, the overal trend of its head conformation, is excessively domestic in nature.
Thirdly, the slope of the forehead to the bridge of the nose. This component gives the characteristic head shape already described for the slim type. Taken as a whole, the typical profile line of the head and face of the slim type is surprisingly reminiscent of the ideal image depicted in Egyptian sculpture (see Fig. 23 and 24). In thestocky type animals this part often shows the opposite tendency, which leads to the more or less prominent nose and brow shape with its distinctive “stop.” We see this, to variable degrees, in specimens of Felis silvestris. With regard to this feature, the same thing applies to our stockily built Shorthair as it does to out German Longhair: the forehead either leading smoothly into the bridge of the nose or being slightly stepped i.e. not descending steeply in the manner of Persian Longhairs.
Fourthly, the hair character. In this case, the hair of slender form of the English cats is "shaved" [very short] to velvety hair, and this is statement is provided for all kinds of short hair. The principles applicable to the stocky form must also be considered in relation to colour and markings.
Here mention must be made of two form factors, one of which seems to be the slender form and the other secondary factor can adversely affect it. It is possible to misjudge the head shape in male animals with strongly developed jowls. Figure 26 shows how such a misjudgment is possible and, at the same time, how this characteristic does not change the "basic style" of the head conformation in the slender form of cat. On the other hand, in some, but not all cases, castration destroys the development of the jowls. The castrated domestic cat usually has a stately appearance and more lustrous colours and, if castration is done while the cat is still growing, it results in increased size. These are all favourable changes which meant that in earlier shows where the castrated cats competed freely, their improved qualities put the breed in danger [note: Schwangart implies that the greater success of neuters became an incentive to castrate the males that were needed to propagate the breed]. In exceptional cases, these benefits are accompanied by the disadvantage of obesity, especially on the torso. This has a destructive effect on the shape of the slender type cats. Figure 25 shows a fine slender head juxtaposed with the baggy, deformed trunk of the same animal; its body shape before castration matched that of the head.
We find contrasts, in the overall form as well as in isolated features, in association with all the colours and colour-groups and with both tabby patterns. I have associated the stocky form with the tiger pattern, and the slender form with all solid colours and colour combinations, although I do not acknowledge markings (tiger and marble patterns) to be the basis of a breed. The reasons for this restriction are partly positive and partly negative in both cases. To begin with the negative, it is simply not feasible to manage two contrasting conformations in all colours and patterns.
Positive reasons for this restriction are:
On the part of the stocky form: the fact that, by its very nature, it goes together with the tabby pattern, particularly the wild colour and related grey colour, and in which animals we often find, as already mentioned, the largest and strongest specimens and those with F. silvestris details such as small ears and an indented brow; add to this the occurrence, as previously mentioned by NEHRING (1888), of intermediate forms between the "sole stripe" of F. ocreata, and the "sole spot" of F. silvestris, which sometimes makes it impossible to distinguish between feral animals and and true wildcats; plus we have seen that the "red nose" (a characteristic that distinguishes F. silvestris from F. ocreata according to E. SCHWARZ) is also characteristic of the gray marble tabby.
Furthermore, I have already predicted the desire to conserve the impact of F. Silvestris in this conformation type, or to intensify and propagate it; as previously stated, the same purpose is served by tolerating the "Acroleucism" attributable to "wild cat flaws" in these greys (white throat, chest patch and belly patch).
[Note: In contrast to the longhair that came to Europe as an Asian luxury cat about 400 years ago, in my opinion our Shorthair cats are purely or substantially of F. Silvestris heritage and still mated with the wild F. silvestris until a few decades ago. Theey may still be doing so in many areas of Asia and Europe. Crossing with this wild type influences the body shape, the nature of the wild colouring and the character of the hair in affected individuals and strains. This can vary widely from case to case as, among other things, it can be very recent. This process makes it possible to associate the appearance of those Shorthair cats resulting from later cross-breeding; they betray the F. Silvestris influence in their conformation and in the domination of the wild colour and grey tiger-tabby in terms of quality and quantity. These days, after the complete extinction of the Nordic wild cat in our country and in view of its rapid decline in the neighbouring countries, it is all the more urgent to introduce breeding measures to replace that natural incursion process.]
Thirdly, I have already given the reasons for the pure-breeding or rather the merit-breeding principle in favour of disciplined selection, isolating the wild-patterned race, again including both wild-coloured and closely related cats.
Fourthly, the desire to bring make the tiger pattern one of the important marking drawing patterns appropriately contrasted with the marble pattern. - Finally, the aesthetic factor of the very attractive appearance of the stocky conformation combined with the tiger pattern, whereas the stocky conformation is inconsistent with the marble-tabby pattern and looks awkward on solid coloured cats.
According to this, my "Shorthair" breed is based on combining the features of the Shorthair with the tiger pattern described above (Figs. 20 to 22). Pattern and background colour also apply in all recognised shades. The hair character may be "strawy", considering the desired impact of Felis silvestris, on the wild-coloured and "slate-grey" cats especially. It also harmonizes with the apparent shape of the animals and goes together with a somewhat bushy tail.
The opposite form, the slender form allowed only in Shorthairs, forms my "slender race" (Figs. 23 to 26) and is limited to all variations of plain domesticated colouring; hence tiger and marble patterns are not allowed. — The positive rationale for this association and restriction are mainly highlighted by the association of characteristics that define the tiger-tabby Shorthair breed. Just as the condition of the unpatterned domesticated colour is a derivative, the slender form was apparently already preferred in ancient Egypt. Natural harmony between pattern and shape is demonstrated by the fact that some of the single-colour cats, most conspicuously the black, and then the blue, showed a tendency towards the slender shape. We give the same reasons for excluding the marble pattern from the slender race as for keeping it independent of the tiger-tabby breed. The section of this work devoted to it demonstrates its multiple claims to breed significance. Its effect is less impaired by the pronounced slender shape of its wearer than it is by the shape of the tiger tabby Shorthairs.
The repeatedly mentioned "smoke cats", which sit between the tiger and marble patterns and the unpatterned cats, create a headache regarding their position relative to the patterned [and therefore stocky] breeds on the one hand, and the slender [and therefore non-tabby] breeds on the other hand. I have decided consider them as belonging to the slender race if the pattern is wholly absent or if is only slightly recognizable on the legs. We have to make the same concession with the Isabelline and cream colours. If the trunk pattern is pronounced in animals where the hair is bright at the base and smoky at the end, they can lay claim to being the tiger-tabby breed or the marble-tabby breed, such as "smoky marble" or "smoky tiger". Raising forms that are intermediate in degree and type debases the pattern.
I alsol introduced features of form in the constitution of the third of my Shorthair breeds, the short-hair marble-tabby (Fig. 27). Like the shorthaired tiger-tabby, it has just one type of marking, the marble-tabby pattern. This combination has a more arbitrary character than the corresponding combination of short-haired tiger-tabby and slender race. In fact, aesthetic reasons were the main reasons here. For this reason, this form may be subjected to the common objection "matter of taste"; however, I have been able to observe that it appeals to taste. There can be no objection to the pattern’s value as a basis for a breed. I believe that I have demonstrated this in the sections devoted to this work.
In my eyes, extremes of conformation are detrimental to the marble tabby pattern. I want a stately, but not really heavily built wear of this pattern. The head should be carried upright, not low as in the thick-necked shorthaired tiger tabby. So on the whole, a proud appearance. The skull should not be excessively slim or broad, the face should be moderately long, but the snout should not be pointed. There is a type of face among Shorthair cats that I would like to cultivate on the marble tabby. I also first noticed this in the marble tabbies, however I am undecided on whether it is really found more often with the marble pattern, or whether it is simply more harmonious with marble tabby than with solid colours. The frequent recurrence of the combination of features that make up this type of face (regularly or at least frequently) makes its unity in inheritance likely. A more difficult but well-established task of livestock breeding is to settle such matters.
It is the so-called "grand face", an expression that becomes readily comprehensible when you look at Fig. 27. Large, wide-open looking, but not fixed or staring, clear eyes are necessary for this.
As already mentioned, marble-tabbies occur in the same colour variants as Shorthaired tiger tabbies.
For the constitution of the three Shorthair breeds I would first of all remind you of my remarks about wild colour, wild patterns and marble patterns, according to which, most likely, the tiger pattern, and then the marble pattern, and it is especially advantageous in both types to have the wild colour as well as grey and deep brown tones. With care, we could even use references to psychological features associated with such characteristics (aptitudes, character). The body shape now vies with pattern and colour in terms of physical and psychological relationships here. In my experience large, stout animals tend to be steady, so to speak, and have prudent behavior. On the whole, we are just beginning to explore these complicated relationships. I consider it proven that constitutional difficulties are associated with certain types of unpatterned domesticated colouring. Then, of course, animals of the Slender Breed also differ. There is no reason to regard the body shape of that race as degenerate.
I have already pointed out that breeding for these three types, tiger-tabby, marble-tabby and Slender Breed, is only advisable in Shorthairs. In the Longhair each such attempt would result in losing the Persian’s highly bred form. In addition, the markings are not nearly as good on Longhairs as they are on as the Shorthair. The second long-haired breed, the German Longhaired which I created, has been described, as pointed out, in terms of on its conformation. These are the same for all colours and patterns, in that they correspond to those of the Shorthair tiger-tabby (with the obvious exception of long hair). As I have already said, it would be "impossible" to have the "slender form" with long hair.
Selective breeding has only been carried out in Persians and Siamese cats - their "family trees" can be seen alongside those of the best pedigree dogs - however we are still dependent on selected individuals in the German Longhair, tiger-tabby Shorthair, marble-tabby Shorthair and the Slender Breed. In this case, we must exercise the significant influence of careful inbreeding, in the sense of C. KRONACHER (1924) and B. LÖHNER (1924).
A look at the Shorthair material which cannot be accommodated in the three Shorthair breeds just discussed, shows us certain combinations that have to be eliminated from breeding based on these principles. These are the tiger and marble patterned Slender Breed cats, the heavy and stocky marble-tabbies, the crossbreeds of tiger and marble, both markings and shape features, and the piebald tiger- and marble-tabbies.
More difficult is the case of the solid colours and bicolours in the medium and heavy build cats. All colour variants are allowable in the slim breed. The majority of such colour cats (a simple expression for such animals) do not naturally belong in that breed. To exclude all of those non-compliant animals from breeding, just to meet the rules on colour, would be a huge mistake and a move that is not fully justified. It is worth restricting the introduction of colours from Shorthairs into the Slender Breed for this to create a similar imbalance as that created in Longhairs by accepting only the Persian type, something I wish to remedy with my German Longhair. The principle also extends to preventiing of further breed development. The recognized breed type must be strictly maintained by breeders, but the breeding system itself must be flexible.
Simply dividing the remaining population of coloured Shorthair cats into other "breeds", for example, basing the breeds on individual colours and colour combinations, would of course be a breed-destroying start from the point of view of my system. I have chosen a different method with the aim of controlling breeding stock and spreading general good qualities (pleasant appearances, benefits of pure breeding, hair quality, colour) in it. I have divided it into colour types and although I have not given them breed status they are eligible for their own breed classes. Premium animals from these "classes" should always be ranked behind those of the recognized breeds, so that a second prize black cat of the Slender Breed ranks above a heavily built black tortoiseshell, etc., coloured cat awarded the same rank. This also increases the likelihood of discovering breedable characteristics that have escaped us, and those those that are still under development. With regard to form, I demand a harmonious impression of these non-breed animals, just I do the representatives of breed classes. On this point I refer back to my criticism of the English so-called "intermediate form".
An example of the current approach to breed formation among coloured short-haired cats which do not belong to the Slender Breeds, is that of the so-called “Belgian”. They are extremely stocky single-colour or bicolour animals and, in most cases known to me, are two-colour piebalds in red- (deep orange-) and- white or black-and-white (also in tiger [mackerel] and marble [classic] and white). The rationale for this variety may be an attempt at selection for a particular use, which will be discussed later, but it is just as likely to be directly influenced by climatic conditions, permanent residence in the ports, docks and ships, or its predominantly fish diet. The main distribution seems to correspond to the field of activity of those reputable ratters in the Belgian port and canal business. In these Belgian cats, the shape in detail does not need to match that of my similarly stocky Shorthair tiger [mackerel tabby]. In particular, you often find in them a combination of a broad head and bridge of nose, proportionate to the face, and a short but pointed muzzle. They are often seen at cat shows. You cannot regard it as an actual race. Similar examples occur even in our breed.
VII. THE OVERALL SYSTEM.
The "Standard of Points" (standard), which I have prepared for the purpose of judging at shows and also as simple guidance for my overall breed classification system (SCHWANGART 1929b) is based on a scale of points just like all other foreign systems and their domestic imitators. The reasons for and against this method of assessment (there is ongoing controversy among various various animal breeding parties) are set out in its Guidelines. There you will also find some other technical basics as well as details on the characteristics of all subgroups of breeds and other breeding classes. The scores for individual features (body shape, head, eyes, hair, pattern, colour, tail, and additional points if necessary) may differ greatly depending on circumstances, but the total number of 100 points is maintained each time, unlike English standards. For example, when assessing a wild-colooured tiger tabby in the Shorthair breed class, out of the available 100 points pattern and colour are worth 15 points, in the silver tiger they are worth 25 points, in the red tiger-tabby 20 points, in all marble-tabbies they are worth 30 points, and in the slender race the head and body conformation are so improtant that together they are worth 60 points. In order to help breed this important unity of form, on the Slender Breed only I allow a predominantly white piebald pattern.
It can not be my intention to reproduce the Standard of points here. However, in order to illustrate what I have said about breed image based on physical characteristics and my concept of breed, I recommend a simple comparison of various officially used classifications
1. the English classification common today (see "Domestic and fancy cats" 1921);
2. the classification of the German "Federation for Cat Breeding" of 1927, at the beginning of my study of the subject, quoted in the 1927 exhibition catalogue;
3. my current classification recognized by the same body.
1. English Classification System.
[Translator’s Note: the letter "j" is not used.]
A. Longhair or Persian cats.
a) White.
b) Black.
c) Blue.
d) Chinchillas.
e) Silbertabbies.
f) Orange and red tabbies.
g) Self-colour.
h) Cream.
i) Tortoiseshell.
k) Tortoiseshell and white.
l) Brown tabbies.
(Under "Tabbies" there is no distinction between tiger and marble patterns.)
B. Shorthair Cats.
a) White.
b) Black.
c) Cream.
d) Orange.
e) Silver, orange, red and brown tabbies.
f) Tortoiseshell.
g) Tortoiseshell and white.
h) two-colour checks.
i) "Manx" (tailless),
k) Siamese.
l) Russian Blue,
m) British Blue,
n) Abyssinian.
(Under "tabbies" Tiger and Marbles are classed together. The "Manx" = severe degenerative loss of tail (refer to the beginning of SCHWANGART and GRAU 1981.)
2. Breeding classification in the Federal for Cat Breeding Catalogue for the Dresden Cat Show at the end of 1927.
A. Long-haired cats.
a) Angora.
b) Blue Persians.
B. Short-haired cats.
a) Siamese Temple cat or Royal cat.
b) Shorthaired tiger-tabby
c) Shorthair solid colour cats.
d) Marble-tabby Cats.
e) Mask cats [Note: bicolour with "tuxedo"pattern].
f) Piebalds.
g) Interestingly marked cats.
h) Half-angoras. [Note: Semi-longhairs]
i) Nubians.
The Shorthair Tiger Tabby breed appeared at this exhibition for the first time. Shortly before that I had published my first reference to its formal founding (SCHWANGART 1927). On that occasion, I had already described the slim breed, but I did not introduce the name until a year later (initially in the non-rigous wording "slim form"). In the 1927 publication I named such animals "Abyssinians" ignorant of the application of this term to a particular English pure breed (see the section above), which appeared as "Nubians" in our country. An attractive name would probably stand the slender race in good steaad. I hesitate to suggest the name "Egyptians". The "marble cats", here in the sense of those displaying any shape of marble [classic tabby] pattern, appeared for the first time under this name applied by me. Traditionally they are called "wheel-cats".
3. Outline of my current breed classification, which has applied since 1929 (publication of my Standard of Points) at Federal cat shows.
A. Long haired cats (Angoras).
Breed 1. Persian. - Subgrouped according to pattern and colour.
Breed 2. German Longhair. – Subgrouped the same way.
B. Short haired Cats
a) Breeds:
1. Siamese cats.
2. Shorthair Tiger-tabby. – Subgrouped by colouor.
3. Shorthair marble-tabby. - The same thing.
4. Slender Race.
b) Breeding classes without breed acceptance:
1. Smoke cats and pearl cats.
2. Medium and heavy solid-colour and piebalds. With subgroups.
3. Stump-tail cats.
C. Dwarf cats (long or short hair).
D. Category for novel types.
E. Representatives of the wild species related to domestic cats.
F. Representatives of other wild small-cat species.
The “Birman” breed would be included under “Longhairs” here; Smoke cats, which correspond to the slender form, will be allowed under this. Stubby tails must have at least 9 caudal vertebrae (refer to SCHWANGART and GRAU). Section D reveals the elasticity of the classification. Representatives of E and F appear at many cat shows. The claim to be made with E. is that such animals are tame. The appearance of F. silvestris is important for inclusion in the shorthaired tiger-tabby breed. C to F are evaluated without using a scale of points.
VIII. MENTAL AND PHYSIOLOGICAL FACTORS. MERITS AND BREEDING LINES..
The starting point of my previous reflections on breed development and my contributions to it in this work was the morphological principle (represented in body shape, colouring, markings). However, breed development also takes place on other criteria, directly physiological and psychological. The fact that a fundamentally morphological viewpoint naturally incorporates these principles has also been shown by previous considerations. They become fundamental in animal breeding where the efficiency standpoint is a decisive factor. This happens above all in actual utility-breeding, which, as recent examples teach, often prevents a cult of form. In our particular case, as an adversary to cat-hostility tendencies and so as not to be misunderstood with my breeding classification, I have several times warned publishers against "presenting the cat as a mere luxury animal". [See note].
[Note: Due to their utility value, the Shorthair population must remain largely deprived of breeding for form for the time being. Should it turn out that morphological features coincide with utility advantages, things may change. The deep differences between dogs and cats will always keep its meaning, that we have an uncultured mongrels [lit: "promenade mixture"], but in most cases it is also aesthetically pleasing.]
In particular the psychological factor is foremost where the animal’s "performance" is desirable in the mind of the human being and to ensure beneficial contact between the two. This corresponds to an animal in its role as a "house pet" ["Heimtier"] a name I have coined for this aspect of pet life. Dogs and cats are regular house pets [See note]). Of course, we want physical fitness in pets, but character and aptitude characteristics are crucial for them. They are already taken into account by the simple individual selection of an animal housemates, but they have surely always contributed to breed development. Reports like that of J. W. AMSCHLER after N. DERGUNOFF (1928) prove that the study of animal character already has scientifically based methods for selecting for desirable traits
[Note: I have repeatedly given warnings on the eminent cultural value of pet ownership, especially at the present time, and on the particular significance and suitability of cats in this regard, (inter alia, in a memorandum "On the Nature and Value of the Cat", 1930, the contents of which are to be evaluated in a different context; see also SCHWANGART 1931).]
The utility value of the domestic cat in protecting the economy and public hygiene is gravely underestimated. Especially since there is a corresponding overestimation of the economic importance of small birds and the idea that the cat is a destroyer of this bird population has become an article of faith and, mixed with this dogma, a ruthless propaganda tries to alienate this remarkably skilled controller of rats and mice.
I have repeatedly criticized this situation and refer to the abovementioned memorandum by myself and to my "Guidelines for the Management of Cat Husbandry" (1931), which aim to improve cat ownership and fully account for the legitimate part of the complaints. In principle, it is important to note that in addition to the advocacy of biologists, we have hygienists in favour of the domestic cat. According to FERD. HUEPPE: "If we had no cats in Germany, we would have to introduce them as soon as possible. But we must also vigorously protect cats against our prejudices and mistaken attitudes in favour of other creatures as our hygienic helpers in public health." The leading French hygienist A. LOIR (1931), Vice President of the International League for Rat Control for 30 years, holds the domestic cat as a protector of domestic and agricultural, storage and warehousing, port and ship operations among other things, as "indispensable" to technical rat control methods. Likewise, in 1928 the International Rat Control Congress recognized the cat as necessary to supplement technical control. It also appealed to ROBERT KOCH's view. In the light of these views I would summarize the advantages of the domestic cat against any kind of technical control: "Activity, constant effectiveness, simplicity of procedure, deterrent effect by mere presence". For deficiencies in technical control, in addition to the evidence opposing its advantages, there are repeated findings such as those of the Munich State Plant Protection Institute (see SELL, 1927) that during rat eradication "the more cautious rats are often prevented from taking the bait by seeing the effects of poisoning on the others," or the census results of A. LOIR (1931), according to which, in the area of port facilities, the overwhelming percentage of trapped rats were male; the same trial should be used at other rat-infested sites and in each of the rat species (domestic rat and brown rat).
The performance of cats in controlling rats and mice is therefore topical. Enterprises involved in the production, storage and sale of food have the same interest in cats as agriculture, shipping has the same interest in it as warehousing, the small household the same interest as luxury trades. Regarding the extermination of mice in homes, forest and field, this is now also seen as a simple matter for breeders. The majority of short-haired domestic cats (including the Siamese) are such good mousers that selection for this ability is unnecessary. We concede that the Longhair is less good due to being bred for luxury.
On the other hand, it is known of the domestic ratter that a not inconsiderable number of Shorthair cats are reluctant to attack rats (although feline ratters are superior to canine ratters in their success. Experiments by LOIR, according to which, in my opinion, cooperation between the two could be considered, and would indeed be possible). This case prompted LOIR to draw up a plan for the selection of ratting breeds and also to make all the arrangements for practical implementation (see A. LOIR, 1931a). Because of his perceptions, especially in Belgian port cities, where cats excel in the rat-exterminating (see above about the "Belgians"), he came to the conclusion that the main requirement was a massive building, and that it was important to enhance and spread this property through breeding. To this end, he advocated concentrating the breeding at a few official posts within the municipalities, and he created an example station of this kind at his headquarters in Le Havre.
Meanwhile, however, the opinion of the preconditions began to change. I myself, after my perceptions, came to the opinion that the mental disposition determines ratting proficiency, not physique. Even daintier cats are quite capable of fighting with rats, weasels, ermine, polecats, whereas some larger, heavier cats do not have the temperament for it. The Paris Exhibition of 1931, where I judged, also proved me right. Among the rat killers exhibited by a large magazine you could see representatives of our Slender Breed as well as massively built animals. In the meantime, LOIR himself has bred a particularly keen strain of ratters out of the Slender Breed. Further, his recent experience in this area of expertise has shown that, in any case, a high percentage of cats than assumed, were keen ratters. Thereafter, it may also be assumed that the cumbersome method of a generally centralized breeding plan (with the requirement that all available cats must reside at one of the official breeding stations) may be disregarded. Where feline ratters are necessary, I find, one usually knows the most suitable animals. In the poorer quarters, where technical control most often fails, people lend each other such cats. This sort of selection is, therefore, warmly advocated, but on the whole it should be left to the interested parties. A leaflet would have to encourage them. For special cases of buying ratting cats (for larger facilities of all kinds), however, LOIR's system is recommended, whether state or private breeding institutions acted in this. The possibility of abandoning a more complicated apparatus is all the more appreciable in this case, since "simple procedure" is one of the advantages of the domestic cat over artificial methods of rat control.
A downside of using domestic cats against rats is not the existence of a minority of unalterably rat-shy cats, but is due to the persecution of cats. Breeding for keen ratters, of course, like any change in breeding direction, can be undone by the circumstance that after seeing a domestic cat crossing the threshold of the home, you can never rely on it being unmolested, or even returning home. Added to this is the circumstance raised by LOIR, which I can confirm that domestic cats, when subjected to persecution, lose the bravado necessary for rat-catching. (One recognizes such animals already as most stick to our "racing pace", as soon as they have to move among humans outdoors). The profession of rat extermination requires the cat to have a certain amount of "self-esteem," and this is lost if it is not protected. LOIR has taken cats that were extremely depressed and restored them to their previous efficient rat-catching state by appropriate treatment of these rat-shy animals.
The principle of the flexibility of my breed classification, which has been repeatedly stated in this paper, paves the way to include the ratter, like any other desired utility-breed, in the cat-breeding business. Should we now allow ratting cats at cat shows? Should we present awards there? This question, of following a plan for utility breeding first became acute in France and there was an immediate uncertainty of judgment. In the 1930 Paris exhibition of the "Societe centrale feline de France," the English breeder H. C. BROOKE (who was amazed on several occasions) judged the Shorthair cats, and passed over the specimens of A. LOIR‘s ratting cat breed as "not breed-like." At the 1931 show, under my judicial office, the management followed my suggestion to award a large encouragement prize to the best ratting cat exhibited by a large magazine. (In addition, one of those cats earned a high score in the Slender Breed class, this award having nothing to do with the exhibitor’s intention.) My proposal was based on the idea that at present recognition, which would cause other companies to follow the example, is highly desirable. Basically, and as a permanent institution, I can not support the showing of ratting cats for the simple reason that ratting tests are not possible in the show hall. The cat’s nature cat complicates the usual kinds of test (as A. LOIR shows with examples) usual for ratters. Nevertheless, I want to make awards possible and the only way I can see to do something similar is they way I have exhibited mine in training tests. Ratting tests must, therefore, take place in the house or property of the owners. They should be set up in such a way that the examinee does not see anything new and unfamiliar in the event. I will return to the significance and the peculiarities of the domestic cat as a trainable subject.
The goal of this breeding direction is to increase and spread the ability and drive to catch rat among domestic cats for their more extensive use in the eradication of other pests. I have already mentioned the propensity and suitability of some cats for destroying even well-armed ones such as weasels, ermines, polecats, and by expelling these and keeping them away cats make themselves useful e.g. on poultry farms. I recommend efforts are made to selectively breed for this ability. — I have also received reliable reports on the extermination of rats by cats. Cases of cats successfully controlling of this elusive pest seem to be commonplace. In the old-school assumption that cats are simply "good for nothing" outdoors, we have failed to pay attention to what they are doing there.
As to the case of breeding ratting cats and the closely related and further-reaching topic of breeding domestic cats for utility, its various diverse mental abilities make this house pet less mentally standardised than it is physically standardised, pointing to numerous courses and possibilities.
A particular mental fitness makes the domestic cat worth farming for its fur, something otherwise based on physical qualities i.e. pelt quality. This is due to the pronounced tendency of most mother cats to adopt the young of others (even the young of prey animals). This gift could also be strengthened by selection. Fur-farming literature contains a number of notable publications on this usage, which has become very popular. (A picture referring to this adoptive relationship "Jungmarder mit Katzen" [Young Marten with Cats] in H. PRELL 1927). By the way, unfortunately there has beean a huge emergence of cat skins from illegal methods as the cat is rarely the direct subject of the fur-farmer. Here I confine myself to a letter containing the expert opinion of a leading informant, Mr. P. SCHÖPS, Managing Director of the Reich Center for Fur and Animal Research, culminating in the sentence: “It is not worthwhile in any way to keep or breed cats for their coat.” (7. VIII. 1931). Personally, I take the view that pet animals, such as dogs and cats, are already basically exempt from the fur production trade. A combination of the resulting use and treatment [of the cat as a fur animal] with the perception and treatment of the same species and even the same individual as a friendly home companion creates a state of mind with "double standards" in the owner, thus promoting hypocrisy and brutality. The lack of value of domestic cat fur to the fur breeding trade coincides with my own opinion is, for me, a stroke of luck.
Of the many favourable instinctive variants known in the domestic cat is the frequent tendency to display its prey to the owner, a procedure that is often practiced very regularly (this facilitates the desired inspection on the type of booty), some have the lesser known peculiarity of bringing home unharmed prey (already spoken of by BREHM), and some cat owners probably know from their own experience that this instinctive tendency can be developed to the spontaneous, and then regular, retrieval of thrown objects. There is a wide range of individual differences, from inability right through to perfection, with regard to walking to heel and free accompaniment, coming when called or to a whistle, the type of devotion, of which there is one towards the home, one generally to humans, one strictly directed towards the person, and there are also transitions between these.
In order to increase desirable qualities, selection and training must work together. It is natural for the dog owner to train his dog efficiently during its adolescence. However, the deceased master-trainer PERZINA said that started teaching his kittens according to their personality during the first weeks of its life, and his tomcat Luchs, an outstandingly gifted individual, proved this by mastering 16 more or less complicated tricks at the age of a few months, astonishing the cat owners who heard about it.
Regarding the degree of trainability, domestic cats show a particularly wide range in their individual psychological sophistication, and the following reminder corresponding to selective breeding seems irrefutable. A connection between trainability and body shape is so far only established in that Longhairs tends to learn more slowly and he compensates for this disadvantage by being more tranquil.
How favourable the general prognosis is, is evident from the statements of the expert, such as the trainer W. VAN PEE (1927): "As a training subject, the cat is harder to work with than the dog, but properly treated and trained, it is far more reliable." (The cases of cat training mentioned here are always humane methods). Naturally, the "correct treatment" of cats differs somewhat from those appropriate to dogs. After all, I don’t hesitate to return to the original interpretation of ancient Egyptian art representating "hunting cats", which was abandoned as a result of the prevailing contempt for cats. It is unclear why the descendants of the easily domesticated wildcat should be lagging behind other species of felids (caracal, cheetah) in theis respect [see note]. An interpretation of the well-known hunting scene also reproduced by W. WRESZINSKI (1 c), where the cat accompanies its masters who are hunting with throwing-sticks and at the same time seizes three apparently injured marsh birds (a hunting yarn! [Jägerlatein!]), they "compete in their hunting zeal with their masters," probably fits the wishful thinking of a hunter of that time as much as it fits one of today.
[Translator’s Note: The caracal and the Indian cheetah (also known as the "hunting leopard" have been used as hunting cats by humans. Some Indian rulers had extensive "stables" of cheetahs – all wild-born as they did not breed in captivity - that were used for hunting and coursing deer.]
To fully understand what I have written about the special arrangements for examining trained cats, which are conditioned by peculiarities of temperament, with certain difficulties being confronted by the preference for consistent training praised by PEE, I would like to add: The cat (which is normally so "absorbed") is easily distracted in an unfamiliar, humanly "prepared" scene. This peculiarity easily leads to the failure of particularly gifted, but also more sensitive, specimens, and in ignoring this circumstance dullness and boldness are rewarded rather than performance, and as a rule, peak performance is missed by the judge. This peculiarity is essentially based on the pronounced orientation need of the settled (familial) loner along with its curiosity and self-consciousness.
As a complement to the examples of enhancement properties mentioned in the section on performance breeding, there are also properties those whose suppression or reduction by breeding is desired.
The tomcat habit of spreading bad smells and causing propery damage by spraying is a considerable obstacle to pure breeding with my form-breeding plan in my racially valuable, but initially, of course, unprofitable new breeds of Shorthair tiger-tabby, Shorthair marble-tabby and Slender Breed. I have discussed some of this in the treatise by SCHWANGART and GRAU (1931). The detrimental effect of breeding is the fact that the majority of newly established breeds are quickly abandoned (either voluntarily or as a result of objections by relatives or residents against keeping uncastrated tomcats) and that the very best and most well-kept cats are mostly castrated males [see grade]. Every cat show convinces me of this second mentioned fact.
[Note: In the above-mentioned work I reported several cases (weak and rare) of female cats spraying, and I recently found a castrated male that sprayed. The secretion, which was only droplets, had only a weak odour, but did harm to paintwork and polishes. The cat was just over 1 year old when he was neutered, the spraying started after about another year.]
A method initially considered against this nasty situation involved finding a procedure for eliminating the "anal sac" used during "spraying" without castration (according to a number of authors [e.g. OTTO KRÖLLING 1927]). According to anatomical findings, this effect should be achievable in the cat by destroying the tissue supporting this function rather than eliminating the sac itself. But even the mere destruction encounters difficulties. My colleague HANS RlCHTER-Dorpat told me (1930) about the status of preliminary investigations on the practical question: “Extirpation of the sac is likely to be difficult to perform, since it is very thin-walled and the external anal sphincter (striated muscle fibers!) adjoins the wall, so that it might not be removed without injury, which will hinder the anal closure. Thus, only cauterisation can be considered, which still easily leads to abscess formation. Perhaps the spraying ability can be suppressed by expanding the narrow excretory duct?”
It is conceivable that selective breeding offers a second remedy against the bad habit of "spraying." Just as domestication has blurred a set breeding season (which occurs in females and they continue to thrive), this could be extended to spraying until there is a permanent change; a drying up due to selection until it stops completely without disadvantaging the tomcats as breeding animals. For example, they are accepted by female cats just as well as other males, indicating there are many important factors in the personal sexual likes and dislikes that characterise this animal (in both sexes). However, a selection for this deficit would be very cumbersome due to the relatively small number of non-spraying tomcats and it is hardly connected to breeding for conformation.
In the meantime it would be crucial if a breeders' association resolved to take the inspired action of keeping an exemplary stud of my short hair breeds. How much should be done by such a stud cat, I leave such details to G. HAAK (1926), according to whom, if necessary, one stud is sufficient to cover 15 to 20 female cats.
Next after this example of a technically "physiological" bad habit I choose the mostly annoying, and preventable issue, of the excess length of the claws in domestic cats. Their development differs considerably. They may become disproportionate to the claw sheath so that they are no longer retractable. It is also conceivable that the defect is due to slackness of the retracting ligaments. (For the mechanics of this see H. VIRCHOW 1926). I once saw partial development of this anomaly where the disproportional length was limited to the hind claws. For personal interaction in the home and for the sake of protecting the environment, we prefer animals with weakly developed claws, but for utility purposes such as rat control, perhaps prefer the other extreme. If the variation is hereditary, as my own experiences have found, nothing stands in the way of selecting from natural stock.
An example of eradicating the drive and tendency to catch birds. The explorer Prof. OSKAR NEUMANN, Berlin, once discussed with me how the small cat species could be distinguished as mouse-catchers, bird-catcher and fishing cats. (The category of "mouse catchers" included the beloved reptile catchers and those that catch other smaller mammal species). Both wild ancestors of the domestic cat are themselves mousing catchers, and are quite moderate climbers and certainly not "tree animals". With regard to the domestic cat in general, unbiased observation of grazing animals confirms the judgment of J. ZELEBORS (1869), issued six decades ago on the basis of investigations into the solution for Felis silvestris, on the basis of which he recommended the protection of this wildcat. However, O. NEUMANN’s statements about the members of all three groups is also true of the domestic cat: she is inclined to take advantage of the other two other food sources [birds and fish] that lie further afield. And on this point, it varies from individual to individualy, as it is also related to the finer details of their personal food preferences. There are birdlovers among them, but many more cats than generally believed do not catch birds. The obvious conclusion that these differences are related to conformation and equating the bird-catching instinct with heavy physique, would be premature. Already a similar experiment in the case of ratting-cats, who probably had more in their favour, was, as you saw, missed. Selection must first be based on simple experience. "Bird-addicted" individuals should be excluded from breeding.
In the sections just completed, examples have shown that apart from the morphologically prominent characteristics (shape, colour, markings) on which my breeding system is based, in the domestic cat’s case physiological, psychological, and "biological" features (in the narrow sense of the term) could also form a breed, by being assigned to a breed, or even without a connection to a breed. To what extent and in what way, in the present state of cat breeding, should such factors be taken into account? How important are they in comparison to the morphological factors?
It is certain that these also deserve attention, in practice as well as scientifically, and that practical steps must be considered urgent in order to improve performance or eliminate disturbing bad habits. On the other hand, however, the breeding scheme that has just been initiated guards more against the dangers of "polypragmasia" [practical meddling] than one that is already more advanced. In our case, this first of all means: disrupting the already well-founded, partly implemented, and at least partly efficient breeding based on on external characteristics that has already begun.
The middle way offered here is illustrated by my example of breeding ratting cats. Also for the whole of performance breeding (for utility and pet qualities) we recommend the widest possible disclosure and introduction of general principles, in conjunction with the necessary precepts of demeanour and nurture, as provided in my aforementioned "guidelines."
CONCLUSION.
On the whole, this essay has given us diverse foundations for an efficient breeding system, the fundamentals of its structure, scope to expand it while preserving unfamiliar raw material and the necessity of supplementing it with principles other than those based on morphology. It has given hints and details on how to do this, and many specifics as well providing general breeding information and issues alongside an inventory of selected proficiencies, and the biological, physiological, hygienic and psychological factors in breeding as the title emphasizes. The identification of problems, as well as accomplishments, is necessary for scientific and practical breeding.
What a step towards this point is already revealed by the breeding classifications reproduced here for comparison with each other. A significant addition is a comparison between some recognized modern works on animal breeding or pet science in general, in their treatment of the domestic cat. A conspicuous cause of resistance is certainly due more to the age-old contempt for the cat as a pet (probably without premeditation by the author), for which I have already stated causes, than to the apparent lack of sound breeding foundations. A change for better consideration seems to be on the way, and is expressed in a renewed appreciation.
O. ANTONIUS (1922) repeatedly mentions the domestic cat for the purpose of showing domesticated changes in comparison with other species. Regarding the formation of breeds, he says only, "one of the few clearly distinguishable cat breeds" was "characterized by it long and fine hair coat". There are dedicated sections about dogs, cattle, sheep and goats, pigs, horses and donkeys, camels and llamas, vut there is not a dedicated section about the cat. M. HILZHEEVIER, in his studies on pet history (1913), considered the cat separately in detail and his pet book dealt with her, along with ferrets and rabbits, on three sides altogether, compared to 49 sides devoted to the dog. L. ÄDAMETZ (1926) often uses the example of the domestic cat as evidence of domestication phenomena and scientific heredity information. He deals with cattle, horse breeds, donkeys, goats, pigs, and dogs in individual sections. His charts also depict "ancestral and domestication areas of some important pet species": camels, dromedaries, llamas, guinea pigs, rabbits, ferrets, ostriches, canaries and many others, but the domestic cat is missing.— In the case of the large animal breeding works of C. KRONACHER it must be said that it systematically emphasizes agricultural livestock breeding. For this reason, even the dog breeds are only listed as an example. Furthermore, his work is under revision, so that only parts of the current version are from the last few years, while the rest is still a somewhat older edition, specifically including the section on "breeds". So far, the domestic cat has been mentioned occasionally in it, in a discussion of the angora character, size changes due to domestication and certain malformations. - The comprehensive zootechnical and livestock encyclopedia of V. STANG and L. WlRTH (Bd. 6, 1929) gives the domestic cat relatively large space; -, 8 pages compared to 28 pages for the dog (with its incomparably richer breed development) - and it also covers them from the benefit perspective. His statements about cat "breeds" are reproduced according to sources, such as, for example, the the very useful book – from a veterinary viewpoint – but failing on the topic of breeds "Le Chat" by E. LARIEUX and PH. JUMAUD and similar cat books based on ill-founded, even legendary, accounts. It is limited to a few examples and does not follow any system. In connection with this we have the clear, small "Pet Science and Pet Breeding" [Haustierkunde und Haustierzucht] by E. FEIGE. On the two pages about the cat (compared to about 10 about the dog) the recognition of their usefulness contrasts with the opinions on breeding. It is not an orginal one: "The cat breeds itself and knows the ways and means to find love easily, as nocturnal roof concerts prove". Now it is probable that other pets, including pedigree ones that are painstakingly bred, like dogs, also find ways to "breed themselves", often to the annoyance of their breeders. The fact that "roof concerts" are now counteracted by controlling breeding and by castration makes the encouragement of proper breeding and husbandry more likely. In addition, may my work serve as evidence of objectively worthwhile breeding goals!
BIBLIOGRAPHY
ADAMETZ, L., 1904. — The biological and breeding importance of pet colours. - Yearbook. Landw. Plant and Animal Breeding, 2, pg. 1-60.
— 1926. — Textbook of general animal breeding. - Publisher Julius Springer, Berlin.
AHARONI, J., 1930. — The mammals of Palestine. - Z. f. Mammalogy 5, pg. 331—332.
AMSCHLER, J. W., 1928. — Character analysis of fur animals (according to a manuscript by N. DERGUNOFF, Moscow). - Fur farming 4, pg. 111-116.
ANTONIUS, O. 1922. — Basics of a tribal history of pets. - Publisher Gustav Fischer, Jena.
— 1930. — Observations on Solipeds [Solid-hoofed animals] in Schönbrunn. V. Bergzebras, Grevyzebras and Zebroide. - The Zool. Garden 2, pg. 261 278.
— 1931 a. — For a more detailed knowledge of the real Quaggas (Equus quagga quagga Gm.). - The zool. Garden 4 pg. 93-115.
— 1931 b. — Remarks on the animals of the Weidholz-imports. - The zool. Garden 4, pg. 204-222.
— 1931 c. — Discussion by the author "Breeding classification and evaluation regulation" etc. -The Zool. Garden 4, pg. 86.
ARMBRUSTER, L., 1921. — New documents about the oldest pet. - Naturw. Weekly 20, Bd., Pg. 193-197.
BAMBER, R. C., 1927. — Genetics of domestic cats. — Bibliogr. genetica 3, pg. 1-86.
BAUM, HERMANN, 1931. — Functional or activity adaptation of living organisms, excerpt from a Rectorial speech 31. X. 1930. - Research, and progress 7, pg. 225 - 226.
BEAUX, DE O., 1928. — Dissertation on some Italian works. - Z. f. Mammal science 3, pg. 55—59.
BEMMELEN, YAN, J. F., 1931. — Colour Patterns of Mammals. – Verh. d. Zool. Ges. Zool. Num. 5. Suppl. Pg.236 - 251.
BIRULA, A., 1916. — The two new cats in Asia (under “News Itineraries and Diverse Events”)- — Ann. Mus. zool. Ac. sc. 21, pg. 1—3.
BROOKE,H. C.1929a. — The Abyssinian Cat. — Published by the “Abyssinian Cat Club” (Engl.), Taunton
— 1929 b. — A suggestion. — Cat. Gossip 4, pg. 223, ebendort.
— 1929 c. — The unique red short hair. — Cat. Gossip 4, pg. 263.
— 1930. — A true albino cat. — Cat. Gossip 6, pg. 118.
BUNGARTZ, JOH. 1896. — Illustrated Cat Book. — Publisher Paul Parey, Berlin.
CASTLE, W. E., 1919. — Siamese, an albinistic colour Variation in cats. — Amer. Naturalist 53.
Discussion about the ancient Egyptian domestic cat (BRUEGSCH, HARTMANN, HIRTH, RUD. VIRCHOW). — Verh. Berl. Ges. Anthr. Ethn. Urg. 1889.
EGGERT, BR., 1931. — About degeneration of the eye and its environmental condition. - Research, and progress 7, pg. 296.
ESSKÜCHEN, E., 1929. — The colouring of domestic mammals in their origin and its importance for the resilience and efficiency of the same. - Arb. German. Ges. Breeding Customer Heft 42, publishing house M. u. H. Schaper, Hanover.
FEIGE, E., 1929. — Pet customer and pet breeding. - Collection "Science and Education". Publisher Quelle & Meyer, Leipzig.
— 1931. — Siamese cat and Russian rabbit. - Review 35, pg. 474-476.
HAAK, GEORG, 1926. — Cat farm. - Fur farming 2, pg. 14.
HAECKER, Y., 1918. — Evolutionary Features [Properties] Analysis (Phenogenetics). - Publisher Gustav Fischer, Jena.
HECK-HILZHEIMER, 1913. — Brehm's Animal Life 12, pg. 111-128. - Publisher Bibliographic Institute, Leipzig and Vienna.
HlLZHEIMER, M., 1926. — History of Natural Breeds of domestic mammals. - Publisher Walter de Gruyter, Berlin and Leipzig.
— 1913. — Overview of the history of pet research, especially the last thirty years. II. The domestic cat. - Zool. annals 5, pg. 233—254.
— 1928. — The reshaping of the skull shapes of domestic mammals due to domestication (a contribution to the race history of domestic animals). - Magazine for animal breeding and breeding biology 12, pg. 85—118.
— 1931. — On the family history of the domestic dog with special consideration of different dog types. – Z. dog research 1, pg. 3—14.
HUEPPE, FERD., 1931. — About the hygienic importance of cat husbandry.— The sick plant, Monthly Paper of the Sächs. Plant Protection [Pest Management]. 8, pg. 75—76.
JANKOWSKY, W., 1931. — New research on hair colours. - Research, and Progress 7, pg. 242-243.
JÜMAUD, PH., 1926. — Cat Breeds. Second Edition. — Published by Saint Raphael (Var.).
KELLER, O., 1908. — The story of the cat in antiquity. — Mitt. Kais. D. Arch. Inst. Rom 23, pg. 40 - 70.
KLEIN, BR. M., 1930. — Face, body and play of the cat. - The Naturalist 7, pg. 201-207 u. B26 - 831.
— 1931. — From the life of three young cats. - Conservation 12, pg. 326-331.
KRIEG, HANS, 1921. — Investigations on the occurrence of coat drawing in mammals. - year. Ver. Vaterl. Naturk. Württemberg 77, pg. XVII—XXI.
KRÖLLING, O., 1927. — Development, construction and biological significance of the anal pouch glands in the domestic cat. - Z. f. d. ges. Anat., Dept. I, Anatomy and Development 82.
KRÖNING, FR., 1924. — About modifiability of mammalian spotting. - Z. induces. Vererbungsl. 35, pg. 113-138.
KRONACHER, C. 1922/3. — General animal breeding, Dept. 3, 5, 6, 2. Issues. — Verlag Paul Parey.
— 1924/8. — General animal breeding, Dept. 1, 2, 4. 3. Issues. - Publisher Paul Parey, Berlin.
— 1924. — The current state of the inbreeding question. - Special issue from the Magazine for Animal Breeding and Breeding Biology 2, pg. 1 - 48. Verl. Paul Parey, Berlin.
KÜHN and KRÖNING, 1928. — About the inheritance of white markings in the domestic cat. - Z. Breeding customer 3, pg. 449-454.
LARIEUX, E. and JUMAUD, PH., 1926. — The Cat. Breeds, breeding, diseases. - Tablets Edition, Saint-Raphael (Var.).
LOIR, A. et Mme., 1931a. — Cat, - its usefulness. Second Edition. – Published by Balliere and son, Paris.
— 1931b. — The Cat - its usefulness. - "Bulletin of the Ratier Cat Club of Normandy" 1931, Le Havre.
LÖHNER, B., 1924. — To clarify the inbreeding problem. - Z. induct. Abst. And Inheritance. 33.
LÖNNBERG, EINAR, 1922. — En blakatt. - Fauna and Flora, Popular Journal of Biology. 1,922.
LORTET and GAILLARD, 1903. — The mummified fauna of ancient Egypt. - Arch. Mus. hist. nat. Lyon 8.
MORTON, CHR. M., 1921. —Domestic and fancy cats. 4. Printed in “The Bazaar”, London.
MURR, E., 1927. — About the development and the finer construction of tapetum lucidum of the felids. - Z. Cell Research and Micr. Anat. 6, pg. 315-336.
— 1931. — Physiological studies on the so-called luminous eyes in vertebrates. I. The light excitability of the domestic cat at dusk. - Zool. Yearbook, Dept of General Zoology and Physiology of Animals 49, pg. 509-632.
NEHRING, A., 1888. — About domestic and wild cats. - Humboldt, Monthly writing f. d. ges. Naturw. 97, pg. 139-141.
OGNEV, S. J , 1930. — Overview of the Russian wildcats. - Z. f. Mammalogy 5, pg. 48—85.
PEE, W. VAN, 1927. — Training of small fur animals. - Fur farming 3, pg. 104—105.
PFIZENMAYER, E. V., 1929. — Hunting and folk pictures from the Caucasus. - Stuttgart.
POCOCK, R. J., 1907 a. — On domestic cats. — Proc. Zool. Soc. I, pg. 143—168.
— 1907 b. — On Pallas Cat (Manul). — Proc. Zool. Soc. 1907, I, pg. 299.
— 1911. — On Tabby cats and some features in the inheritance of their coat patterns and colour. - The Mendel Journal 1911.
— 1916. — Some dental and cranial variations in the Scottish Wildcat (F. silvestris.) – The Ann. and Mag. Nat. Hist. (11) 18, pg. 272—277.
PRELL, H., 1927., — The mating season of the true martens. - Fur farming 3, pg. 223-228.
PRZIBRAM, H., 1908. — Inheritance test on asymmetric eye colouration in Angora cats. - Arch. For Dev. Mech. 25, pg. 260-265.
RASELLI, AD., 1923. — Morphological and functional information about the muscular system in the iris of the cat. - By Graefe's Arch. For Ophthalmology 111, pg. 310-329.
RENSCH-, B., 1929. — The principle of geographical racial circles and the problem of speciation. - Publisher W. de Gruyter & Co., Berlin.
RICHTER, HANS, 1928. — On the distinction between a Tapetum cellulosum and fibrosum in the eyes of domestic mammals and the formation of the shades of Tapetum lucidum. - Munich. Veterinary Weekly 79.
SATUNIN, K. A., 1904. — The black Wild-Cat of Transcaucasia. — Proc. Zool. Soc. 1904, II, pg. 162—163.
SELL, 1927. — Rat bacilli and immunity of the rats. - "Animal Exchange", Dresden-Freital, unnum.
SCHNEIDER, K.M. — 1930a. — A case of hereditary partial albinism in the Lion. The zoological Garden 2, pg. 274-278.
— 1930 b. — Remarks on species of African animals introduced by Christoph Schulz in 1929. - The Zoological Garden 3, pg. 19-28.
SCHARFF, R. F., 1906. — On the former occurance of the african Wild-Cat in Ireland. — Proc. of the Royal Irish Ac. 26, pg. 1—12.
SCHWANGART, F. 1927. — An Attempt to lay the foundations of cat breeding. – Pages on cat breeding and cat protection (animal exchange). Freital (Sa.) 1927, unnumbered.
— 1928 a. — Personal characteristics in animals and animal standards. - Yearbook of Characterology 5, pg. 101-138.
— 1928 b. — Breeds and typical customers of domestic cats. - Works of the Reich Central Station for Fur Animal Research. 1928.
— 1929 a. — Race history, pedigree and breeding system of domestic cats. - Publisher A. Heber, Leipzig.
— 1928/30. — Notes on cat breeding, from the essays: 3) On the Siamese standard; 4) shape, pattern, colour, its breeding value for our shorthair; 7) Angora, Persian, German, Longhair; 9) Blue Shorthair. — Cats friend (animal exchange, issue B.), in numerous issues.
— 1929b. — Breeding classification and assessment rule (standard) of the Federal Government for cat breeding and cat protection, authoritative as a directive, etc. - Federal Publishing, Freital (Sa.).
— 1931. — On the economic and ethical importance of the domestic cat, and: Guidelines for regulating cat ownership. — The sick plant, Monthly Paper of the Sächs. Plant Protection [Pest Management]. 8, pg. 74—75 and 76—79.
SCHWANGART, F., and GRAU, H., 1931. — About malformations, especially hereditary tail deformities, in the domestic cat . — Journal for Animal breeding and.breeding Biology 22, pg. 203—249.
SCHWARZ, E., 1930. — The wildcat of the Balearic Islands. - Zool. Num. 91, pg. 223-224.
SIMPSON, FR., 1928. — Cats for pleasure and profit. — Published by I. Pitman, London.
STANG, V., und WIRTH, D., 1929. — Veterinary and animal breeding 6, Article "cat".
TELSCHOW, 1911. — Outline for of modern sheep breeding. - Hannover 1911. (Cited after Essküchen).
TJEBBES, K., 1924. — Crosses with Siamese Cats. — Journ. of Genetics 14.
TOLDT, K., 1912. — Epidermal stripes, hair stripes and wild markings of the domestic cat. - Verh. Zool.-bot. Ges. Vienna 62, pg. 16-27.
VOSSELER, J., 1927. — Angora cats and Angora goats in their homeland. - Fur farming pg. 25-27.
VIRCHOW, HANS, 1926. — Mechanics of tiger toes. - in Mammal science 1, pg. 64-80.
WHITTING, P. W., 1919. — Inheritance of White-spotting and other colour characters in cats. Amer. Naturalist 53.
WOLTERECK, R., 1931. — Inheritance Science and Endemic Species. - Research, and Progress 7, pg. 164-165.
WRESZINSKI, W., 1923. — Atlas of ancient Egyptian cultural history. - Leipzig.
ZELEBOR, JOH., 1869. — Wildcat and Imperial Eagle. Albert Hugo's Hunting Journal 7 (reprint from "Der Urwähler, organ of the suburbs of Vienna" 1869).
Description of Figures.
Fig. 4. High-bred Persian (chinchilla). By Frau von Larisch, Wolftitz Sa.). Photo R. Zacharias, Regensburg.
Fig. 5. German Longhair (reddish yellow). By Countess Montgelas. Photo Georg Zschäpitz, Leipzig.
Fig. 6. Birman cat. From the "Revue Feline de France" (Clichy - Seine) 1981.
Fig. 7. Felis silvestris SCHREB. Northern European form. From "Meerwarth and Soffel".
Fig. 8. Wildcat (Felis ocreata haussa THOS). After Otto Antonius from "The Zool. Garden 1931. pg. 204." Phot. Dr. Grögl.
Fig. 9. Felis ornata macrothrix ZUKOWSKY. Original photo, and property of K. Hagenbeck, Stellingen.
Fig. 10. Manul (Felis manul PALL.,) Photo Georg E. F. Scholz, Bln.-Friedenau; from Brehm's Animal Life 12, Table IV Predators, Nr, 3.
Fig. 11. Felis (Catolynx) chaus GÜLD. Original photo, and property of K. Hagenbeck, Stellingen.
Fig. 12. Siamese cats. Cattery of Mrs. E. Sacher-Petri, Wroclaw. Phot. "PhotoKrapp", Wroclaw.
Fig. 13. Blue short-hair with "Persian head". By Mrs. Hansen, Kiel.
Fig. 14. As Fig. 13. A cub.
Fig. 15. So-called "Abyssinian". Breeder H. C. Brooke (Taunton). In the possession of the zoological garden Vienna Schönbrunn. a) cat "Ras Tafari", b) cat "Bun".
Fig. 16. Marble pattern. From the painting "Ulysses at Circe" by Karl Ruthart 1666. Phot, the Dresden State Gallery.
Fig. 17. Implied marble pattern, disrupted blotches. From O. Keller, 1908, p. Literature certificate. Altapulisch.
Fig. 18. Mixture of tiger and marble pattern. Phot. R. Zacharias, Regensburg.
Fig. 19. Transition between tiger and marble pattern. a) and b) the two sides of an animal. Photo. Emmy Schmidt, Kötzschenbroda (Sat.).
Fig. 20. Wild-coloured Shorthair "Silvester". By Ernst Braun, Berlin. Photo. O. K. Vogelsang, Berlin.
Fig. 21. As Fig. 20.
Fig. 22. Shorthair striped tabby "Simson". By J. Lesti, Vienna.
Fig. 23. Slender race. Black cat "Moro". By Privy Councilor Prof. Dr. Medicine. Karl Woermann, Dresden Phot. Studio Ursula Richter, Dresden.
Fig. 24. Slender Breed. Black cat "Maja". By Mrs. Alma Schulze, Dresden Phot. "Dresden photography Workshops".
Fig. 25. Slender Breed. Neuter. By Mrs. Leopoldine Hofer, Vienna.
Fig. 26. Slender Breed. Male cat. By Mrs. Etha Richter, Dorpat. Phot. Studio Ursula Richter, Dresden.
Fig. 27. Shorthair classic tabby breed. Male. Federal Exhibition Berlin 1929.
Completed December 1931
DR FRIEDRICH SCHWANGART “KATZEN-SCHWANGART”
The almost-forgotten Friedrich Schwangart was the “discoverer” and founder of the German Longhair cat and writer of books about cats. He was born in Munich in 1874. When he was seven years old, his parents emigrated to Amsterdam in the Netherlands where he attended school. A few years later they moved to Thuringia, Germany, where he attended middle school. His family’s wanderings meant he attended the Friedrichs-Werder-Gymnasium in Berlin, and later the Maximilians-Gymnasium in Munich.

At his father’s request he studied medicine, but during his life he also studied philosophy, psychology and literature. Around 1899, he focused on the natural sciences and in 1902 he gained a doctorate in zoology. He also studied botany, anthropology, ethnography, ethnology, prehistory, palaeontology and history and was an accomplished poet. In 1902 he married Ellie Wellner and they went on to have 3 children. In 1903 he became a member of the German Zoological Society and for many years he occupied various zoological posts and had an especial interest in insects and arachnoids. Later on his passion for psychology was rekindled and he became interested in animal psychology including comparative pet research. He was especially interested in the domestic cat and their non-domesticated relatives and became a recognized Felidae specialist.
He was opposed to war and in August 1915 he was ordered to serve as a bacteriologist in a military hospital in Dresden, remaining there until May 1917. The war disturbed him and he turned his attention to poetry, his anti-war sentiment being apparent in his verse. In 1921 he suffered a serious accident and a disabling knee injury that plagued him for the rest of his life. For some time he was unable to work and turned to his other interests including membership of the “Society of Friends of the new Russia in Germany.” In 1925 he took over the management of a local group of the “League of Friends of the Soviet Union.” His wife, Ellie, supported his work and was a women's rights activist. This set the couple apart from bourgeoisie society in Dresden.
In 1928 Schwangart became honorary professor at the Technical University in Dresden and was primarily devoted to mammalogy, and was a member of the “German Society for Mammalogy”. In 1933 he was forced to return to Munich. He had been a member of the “League for Human Rights” from an early age and Ellie was a member of the “Women's League for Peace and Freedom”. Both organizations were dissolved in 1933 by the National Socialists and the members were persecuted. Back in Munich Schwangart worked as a lecturer at the Technical University and for the Bavarian Zoological State Collection. His love of cats awoke in those years and he devoted himself to the research of wild cats and domestic cats and was known as “Katzen-Schwangart.”
Schwangart was opposed to National Socialist rule and the Second World War. He remained in the Munich area, but the last two decades of his life are obscure and he rarely published articles. He died in 1958 in Gräfelfing, near Munich.