
EARLY SCIENTIFIC PAPERS ABOUT CATS
1912. DONCASTER L: Sex-limited Inheritance in Cats, And its Bearing on the Sex-Limited Transmission Of Certain Human Abnormalities.
Science, N.S. 36, 144.1912.
(Only the cat-related part is included here)
The difficulties in the way of an elucidation for the human cases arise chiefly from the impossibility of distinguishing a transmitting female except by the fact that she has affected sons. In the hope of throwing further light on the question, I compared my summaries of human pedigrees with data which 1 have collected during several years of colour-inheritance in Cats, and find that they are in most respects so closely similar that I believe the inheritance of certain characters in the Cat may provide a solution of the problems which cannot be answered by means of human pedigrees. In the case of the Cat the “trans¬mitting female" is visibly different from the non-transmitting, and the most serious source of error affecting the human data is thus avoided.
The character in the Cat which appears to correspond in its inheritance with the sex-limited affections in Man is the orange colour as contrasted with the black which corresponds with “normality.” In 1904, in a short note on the subject, I concluded that in the Cat the orange colour is dominant over black in the male, but only partially dominant in the female, so that the female heterozygote is tortoiseshell [Proc. Camb. Phil. Soc. Vol. XIII. p. 35.] The existence of sex-limited inheritance was at that time scarcely known, but I mentioned the fact that among my collection of data there was no case of an orange male mated with a black female giving orange male kittens; the females from such a mating were tortoiseshell, the males black. Subsequent collection of further data has shown con¬clusively that the transmission of the orange colour by the male is sex-limited, and the same result has been arrived at independently by C. C. Little from his own experiments [Science, May 17, 1912].
In general, the results obtained with Cats are as follows:– an orange male mated to a black female gives black male and tortoiseshell female kittens; in the converse cross, orange female by black male, the male kittens are orange, the females tortoiseshell. The orange male thus usually transmits orange to his daughters only, the orange female transmits it to all her offspring of both sexes. A tortoiseshell female by black male gives orange and black males, tortoiseshell and black females, showing that a female heterozygous for orange transmits the orange factor to some kittens of both sexes. The colours cream and blue appear to be dilute forms of orange and black, and to be inherited similarly. In what follows I shall use the term “yellow” to include orange and cream, “black” to include black and blue, tortoiseshell to mean a mixture of yellow and black, and, when necessary, “blue-cream” to describe the dilute tortoiseshell. The data given are derived almost entirely from correspondence with numerous breeders, but include the litters given by Little in the article in ‘Science’ referred to. In the information obtained from breeders I have made every effort to insure that all inaccuracies are eliminated, but it is possible that a few mistakes may have been included. Where I have good cause for doubting the accuracy of a record, it has been omitted from the tables.
A summary of the data collected with regard to the inheritance of the yellow and black colours in Cats is given in the accompanying table. It is noticeable that the divergences from equality of the two sexes are similar in kind to those observed in the case of human sex-limited diseases, where, however, as has been seen, it is doubtful whether they are significant. In the offspring of yellow male mated to black female there is some excess of females (61 female, 50 male), as is also the case among the offspring of men affected with one of the four diseases mentioned previously; among the offspring of tortoiseshell females by black males there is a considerable excess of males (67 male, 35 female), as has generally been found to be the case among the children of women who transmit the diseases.
Another point of great importance is that there is evidence that the sex-limitation of the transmission of the yellow factor by the male is not absolute, for thirteen black or blue females are recorded from the cross yellow male mated to black or blue female, five from yellow male by tortoiseshell female, and three tortoiseshell females from one mating of yellow male by yellow female. It should be mentioned that the majority of the eighteen black or blue females from yellow sires were blues, and it is possible that the simple explanation that sex-limitation is not absolute, is not the true one in every case. Apart from the possibility that the sex may have been wrongly recorded in some instances, it is not completely certain that a young kitten may not sometimes be recorded as a blue when it is in reality a blue-cream. The breeders of whom I have enquired on this point are not agreed; some say that blue-creams are always easily distinguished from blues at birth, others that mistakes may be made. In some cases at least these blues grew up and seem to have been undoubted blues, and I know of no case of two blues mated together giving creams, as should happen if an apparent blue may ever be heterozygous for cream. In any case, the few black females from yellow sires, about which there appears to be no doubt, seem to prove that the sex-limitation of the transmission of yellow by the male is not absolute.
Another point of considerable importance in connexion with the comparison with the human cases is the ratio of the yellow to black males in the offspring of tortoiseshell female x black male. As was said above, in the offspring of transmitting females in the human cases, it is generally believed that there is an excess of affected sons, but Lenz regards this as due to the fact that a transmitting woman can only be identified by having at least one affected son, and that this inevitably raises the apparent ratio of affected to unaffected. When only large families are considered, this excess disappears in the case of Colour-¬blindness and Night-blindness, but remains in the Nystagmus and Haemophilia totals, chiefly owing to the very great excess of affecteds in certain pedigrees, but it seems doubtful whether even in these affections it can be regarded as genuine. In the Cat, in which the “transmitting female” (tortoiseshell) is visibly different from the “non-¬transmitting” (black), a small excess of “affected” (yellow) over black is found (35 yellow: 29 black), but as in the human cases there is also an excess of “transmitting” (tortoiseshell) over “non-transmitting” (black) daughters, (21: 12). The numbers are small, and further data are required before they can be regarded as significant, but they give no support to the suggestion discussed above that the excess of affected males is due to partial coupling of the factor for the affection with a sex-factor in the gametes of the transmitting female.
One of the most interesting questions connected with the inheritance of these colours in the Cat is the nature and origin of the rare tortoise¬shell males. Three of these are included in my records, one (a blue-cream) from the mating yellow male with black female, one from tortoise¬shell female by black male, and one from tortoiseshell female by yellow male. Two tortoiseshell males are also recorded from this last mating in a note in ‘Fur and Feather’ for May 10, 1912. I know of no other case in which the male parent was known; the few others that I have met with have been produced by tortoiseshell females by unknown sires. It has been shown above that there is reason for believing that the sex- limitation of the transmission of yellow by the male Cat is not absolute, since a small proportion of black females are produced from yellow sires, and if this is so, there should theoretically be as many cases in which a yellow male transmits yellow to his sons, as there are of his failing to transmit it to his daughters. I know of no satisfactory record of a yellow male mated to a black female having yellow sons, but suggest that the tortoiseshell male is produced when, exceptionally, yellow is transmitted by a yellow male to a son [See also p. 21 below].
Very little is known of how a tortoiseshell male transmits colour-factors to his offspring. The few breeders who possess them commonly mate them to tortoiseshell females, in the belief that this is the most likely mating by which to produce tortoiseshell male offspring, and I know of only one mating with a black female, which is the one required to test the matter thoroughly. In this one case the female was not kept in confinement, so that although the pairing was seen, the parentage of the kittens cannot be regarded as certain; the only recorded kittens were a black male and a tortoiseshell female. When mated with tortoiseshell females, tortoiseshell males appear to behave like yellows, giving yellow and black male, tortoiseshell and yellow female offspring. Sir Claude Alexander, who has made many such matings with one of his well-known specimens, writes of this tortoise¬shell male that with unrelated tortoiseshell females he “sires tortoise¬shells freely,” but that with his own tortoiseshell daughters he gives chiefly yellows, with only an occasional tortoiseshell female or a black. Unfortunately, no record of the sex of these kittens appears to have been kept; no tortoiseshell males were produced.
The facts, then, as far as they are known, of the transmission of the yellow colour in the Cat, may be summarised thus. In the female the factor for yellow when homozygous produces orange (or cream when dilute); when heterozygous produces tortoiseshell. In the male the presence of the yellow factor normally produces orange (or cream), but such males are not homozygous, for in general they transmit the yellow factor to their daughters only. Exceptions to this rule, however, occur, not apparently very rarely, for black females from yellow sires are frequently recorded. Less common are tortoiseshell males, about which little is known, except that they may apparently derive the yellow factor from either parent, and that there is no recorded case of their having tortoiseshell male offspring.
Until more data are collected, it seems of little value to attempt to express these facts in a factorial scheme, for no means of testing such schemes are available. Certain points of importance, however, may be considered. It has been suggested by Little [Science, May 17, 1912] that both the black and yellow factors are sex-limited in the male Cat, and this suggestion is in harmony with the fact that a female heterozygous for yellow is tortoise¬shell, while the heterozygous male is yellow. For if the factor for black were not sex-limited in the male, one would expect the male heterozygous for yellow to be tortoiseshell as in the case of the female. If all males containing the yellow factor were yellow, and if there were no exceptions to the rule that the yellow male transmits yellow only to his daughters, the case of the Cat would fall quite simply into the scheme which I recently suggested for characters which appear to be dominant in one sex and recessive in the other [Journal of Genetics, 1. 1911, p. 377]. This suggestion was that one sex (in the Cat and in Man the female) is normally homozygous for a factor for “normality” (NN), for which the normal male is heterozygous (Nn), and that N is constantly coupled with a sex-determining factor for which the female is homozygous, the male heterozygous. In the Cat the factor for normality would be the black determiner, which may therefore be written B instead of N; then if X is used for the sex-factor, adopting the notation commonly used in America, a black male would be BX.bx, a black female BX.BX. If then the yellow colour be caused by the loss of the factor B, the yellow female would be bB.bX, the tortoiseshell female BX.bX, and the yellow male bX.bx.
This scheme, however, has the obvious defect that it represents the black male and the tortoiseshell female as of the same factorial composi¬tion as far as B and b are concerned (Bb in each case). An improvement might be to represent the yellow factor as a modification of B rather than its absence; if the modified B which gives rise to yellow instead of black be called Y. the yellow female would be YX.YX, the tortoise-shell female BX.YX, and the yellow male YX.bx. Since Y is normally coupled with X, a yellow male would then normally transmit Y only to his daughters, and since when he is mated with a black they would receive B from their mother, they would be tortoiseshell, thus:– Fig. 2.
If now, exceptionally, Y became dissociated from X in the gametes of the yellow male, and were thus transmitted to a son instead of a daughter, the result would be: Fig. 3.

The female offspring would in this case be black, but heterozygous for B instead of homozygous, and the male, since he contains both Y and B, would be tortoiseshell.
I have one record of a tortoiseshell male produced by tortoiseshell female by black male. The kitten died when only a month old, but the breeder is confident that it was a male. If, as suggested, the tortoiseshell male has the constitution YBXx, this could only arise in this instance by the tortoiseshell mother transmitting YX, the black father Bx; the tortoiseshell male would then arise by the exceptional association of B with x in the spermatozoa, instead of B with X [There is an additional piece of evidence supporting the hypothesis that the factor B is sex limited. I have a record of tabbies being produced from yellow male x black female. Tabby is known to be epistatic to black, but presumably can only appear in the presence of the factor B. The yellow male which transmits the tabby factor does so to both sexes, the male kittens which receive the tabby factor being brown tabbies, the females (only one recorded) “tabby-tortoiseshells.” It is clear therefore that the tabby factor is not sex-limited. If, however, the black factor were present in the orange male, it is probable that the cat would be tabby or tabby-and-yellow instead of yellow. If, therefore, a yellow male can in some cases transmit the tabby factor, additional evidence is provided for the assumption that the black factor is not present. Tabby tortoiseshell males, though very rare, do exist, and are presumably tortoiseshell males which bear the tabby factor in addition. One was exhibited at the Crystal Palace Show in 1912.].
The scheme, outlined above is satisfactory up to this point, but it has two disadvantages. It involves the assumption that the occasional black females produced from yellow sires are of different constitution from normal blacks (BX.bX instead of BX.BX), for which there is no direct evidence. The point could be tested by mating such a black female with an orange male; if the female were heterozygous for B, it would then give not only tortoiseshell females YX.BX, but also yellow females of constitution YX.bX. This experiment has never, as far as I know, been made. The second objection is more serious. The tortoiseshell male should on this hypothesis be Yx.BX, and since B is coupled with X. such a male should transmit B to his daughters and Y to his sons. Mated to black he should have black female and tortoise¬shell male offspring; mated to a tortoiseshell he should have tortoiseshell and black daughters, yellow and tortoiseshell sons. Of the mating of a tortoiseshell male with a black I have only one somewhat doubtful record, but hope to be able to test the matter with a tortoiseshell male now in my possession; in the several matings with tortoiseshell females which are reported, no tortoiseshell males were produced – the tortoise¬shell male behaved as far as can be judged from the very incomplete reports as if he were a yellow.
Other possible schemes might be suggested, for example that yellow is due to an additional factor modifying B and usually coupled with it, but occasionally becoming dissociated so that the yellow male would transmit B to a female kitten and the modifying factor to a son. The tortoiseshell male so produced, however, would still receive the modifying factor from the male parent and B from the female, and it would be expected, if these factors followed the ordinary rules of gametic coupling, that they would show repulsion in the gametogenesis of the tortoise¬shell male, so that again the modifying factor would be transmitted to the male offspring and tortoiseshell males would be produced as in the previous scheme.
It seems clear, therefore, that the data at present available are inadequate for the formulation of a factorial scheme of colour-inheritance in Cats, which shall be in accord with previously known cases of sex-limited inheritance and of gametic coupling in other forms. For anyone, however, who is able to undertake the work, the experiments required are fairly clear, and the importance of the case is increased by the fact that the transmission of the yellow colour in the Cat is in close agreement, both in its normal course and in the exceptions which oc
cur, with the transmission in Man of the sex-limited affections Colour¬-blindness, Night-blindness, Nystagmus and Haemophilia. Although so much has been written about the transmission of these diseases, no agreement has been reached, and it seems probable that the inheritance of colour in the Cat, with which definite experiments can be made, and in which the “transmitting female” can be distinguished at sight from the “non-transmitting,” may provide the clue which the most complete human pedigrees would fail to yield.

THE TORTOISESHELL CAT - Phineas W. Whiting, University of Pennsylvania
The American Naturalist, Vol. 49, 1st August, 1915
In The Journal of Genetics (June, 1913), Doncaster has summarized genetic data dealing with the tortoiseshell cat. The records are collected from fancy breeders and from the work of Dr. C. C. Little.
Aside from certain disputed points the inheritance is in accordance with simple sex-linkage and is analogous to the human defects-color-blindness, night-blindness, nystagmus, and hemophilia, and to the thirty or more sex-linked factors of Drosophila.
If the factor for yellow be represented by Y and its allelomorph, the factor for black, by B, the lack of either by b, the sex factor by X, and the allelomorph of X by x, the normal zygotic possibilities are as follows : YX-bx = yellow male. BX-bx = black male. YX-YX = yellow female. BX-BX= black female. YX-BX = tortoiseshell female.
It is obvious then that there can be but two classes of males, while there are three classes of females. Difficulties arise when it is attempted to explain the occurrence of black females produced either by the mating of a black female to a yellow male which should give only tortoiseshell females and black males, or by the mating of a tortoiseshell female to a yellow male, which should give only tortoiseshell and yellow females and black and yellow males. The occurrence of the rare tortoiseshell male is also the cause of considerable difficulty. In one mating out of seventeen of yellow females to yellow males there were produced three tortoiseshell females. There are recorded in addition from the seventeen matings forty yellow females and forty-eight yellow males which are in agreement with expectation.
In order to explain these discrepancies it is suggested that possibly the linkage of Y with X is not absolute. Yellow males may then produce gametes bX and Yx in addition to the normal or more frequent gametes YX and bx. Gamete bX is female determining, while gamete Yx is male determining and yellow bearing. The latter gamete should produce a tortoiseshell male when it meets an egg BX.
On this hypothesis we should expect the tortoiseshell males to be as frequent as the anomalous black females from yellow fathers. Prom the matings recorded there are eighteen anomalous black females and only three tortoiseshell males, and one of these tortoiseshell males had a black father. There is a further objection to this hypothesis inasmuch as it is not explained how gamete bX differs from BX. Doncaster admits these difficulties, stating that further work is necessary before a definite conclusion can be reached.
In a more recent paper [Quarterly Journal of Microscopical Science, February, 1914. ] Doncaster has suggested non-disjunction of the sex-chromosomes in oogenesis as a possible explanation. This explains the matroclinous [inherited from the mother] black females, but fails to account for the lack of an equal number of patroclinous [inherited from the father] yellow males. It also fails to account for the tortoiseshell male and the occurrence of tortoiseshell females among the offspring of yellow by yellow.
In a series of experiments begun upon cats at the University of Pennsylvania during the last year, the tortoiseshell problem has been especially investigated. A yellow Persian male was crossed with common cats-black, maltese and tabby. The results, although not at present extensive, are sufficient to explain, at least in part, the anomalies observed, and to suggest a simple explanation for the occurrence of unexpected classes.
When the yellow male was crossed with a maltese female, a maltese male and two blue and cream females were produced. The blue and cream is the maltese or dilute tortoiseshell. When mated to a black female the yellow male produced both dark and dilute kittens. This shows that the black female was heterozygous for dilution. Two of the males were black and two maltese. The two females were dark tortoiseshell. When the yellow male was crossed with a dark tabby, there were produced dark and light tabbies and maltese. Blacks are also to be expected from this mating. The mother is evidently hybrid between tabby and black and between black and maltese. The female offspring showed yellow : the male offspring were without yellow except for tabby striping*.
The female offspring obtained from these matings may be arranged in a series, ranging from one that is predominantly yellow to one that is maltese except for a few cream-colored hairs. The maltese with a few cream hairs occurred in the litter of three above mentioned, which included also a maltese male and a maltese female with a small cream patch.
It may be readily understood how a maltese cat with a few cream hairs or its intense form, a black cat with a few yellow hairs, would be recorded as maltese or black, and it is reasonable to suppose that further segregation of distribution factors in the direction of black would have produced a fully black female. This may be compared with conditions in the guinea-pig in which yellow spotting is continuous 'with total black. The essential differences are that in the cat we have a factor for yellow allelomorphic to a factor for black, that these allelomorphs are sex-linked, and that either alone is sufficient to produce its expected color, but that when one is balanced against the other, as in the tortoiseshell female, other factors governing the relative amounts of the two colors can act and produce continuous variation from yellow to black.
The three tortoiseshell females from the mating of yellow by yellow may be explained by supposing that the mother was gametically a tortoiseshell plus a sum of yellow extension factors and minus a sum of black extension factors.
The occurrence of the tabby factor brings in a restriction of the black pigmentation producing yellow stripes. It is therefore much more difficult to distinguish a tabby from a tabby-tortoiseshell than a black from a tortoiseshell. "We have had a few tabby-tortoiseshells that would have been recorded as tabbies if close examination had not been made.
Another source of error in records involving the tortoiseshell pattern may be introduced by the occurrence of white spots". Doncaster makes no mention of these in his paper, so that it is possible that they did not occur in the animals recorded. In what is genetically a tortoiseshell and white cat the incidence of the white spotting may happen to be at just those points which would otherwise be yellow.' Thus the occurrence of black and white daughters from yellow males may be explained. It is possible also that the yellow mother of the three tortoiseshell kittens recorded from the mating of yellow by yellow may have been white at points which, if pigmented, would have been black. She would then have been genetically a tortoiseshell and white and some tortoiseshell kittens would have been expected.
I would suggest as a plausible hypothesis that the rare tortoiseshell male is genetically a yellow with an extreme of black extension factors or a black with an extreme of yellow extension factors. This hypothesis is rendered more probable by some slight evidence showing that male tortoiseshells breed like yellows. There is then no need for assuming in the cat either breaks in sex-linkage or non-disjunction of the sex chromosomes in oogenesis.

TRICOLOUR INHERITANCE.III. TORTOISESHELL CATS - Heman L. Ibsen (1916)
University of Wisconsin, Madison, Wisconsin [Received June 6, 1916]
1 Papers from the Department of Experimental Breeding of the Wisconsin Agricultural Experiment Station, No. 9. Published with the approval of the Director.
Doncaster has published several papers dealing with the inheritance of color in cats, the first of which appeared in 1904. Little (1912) and Whiting (1915) have also written on the same subject. Both Doncaster and Little have established that the colors orange (yellow) and black are sex-linked. According to them the female is homozygous and the male heterozygous for sex. On their interpretation a female bearing the orange factor in one chromosome and the black factor in the other is a tortoiseshell, i.e., an animal spotted with black and orange. Black and orange are, accordingly, allelomorphs, but neither is dominant to the other. Orange females are represented by Doncaster (1913) as YX.YX; orange males, YX.bx; black females, BX.BX; black males, BX.br; and tortoiseshell females, BX.YX. Little’s method of repre- senting the above color types is essentially the same as Doncaster’s.
For the normally expected color types Doncaster and Little have the same interpretation, but they differ as to the origin of the unexpected classes (see table 1). Little considers the rare tortoiseshell male a mutation, while Doncaster thinks it may be due to Crossing over in the male. The latter has presented his idea in the following manner:
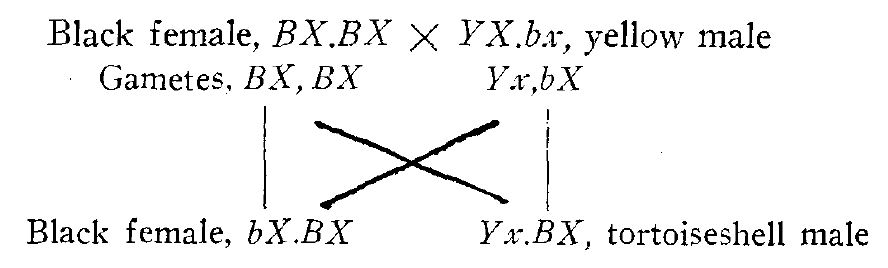
The above scheme would also account for the unexpected class of black females which occurs in the black female X yellow male mating.
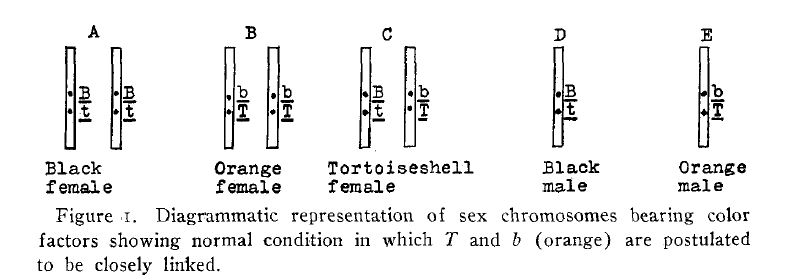
This table gives Doncaster’s (1913) data. The off spring about which there was any doubt are not included. The numbers in heavy type represent the individuals of the normally unexpected classes.
Doncaster admits that there are some difficulties connected with his interpretation. Among these are his bX.BX black females, which as he says should give, when mated to orange males, not only tortoiseshell females but also YX.bX orange females. He fails to point out that they should in addition give bX.bx males. Since b means the absence of both black (B) and orange (F) one would have to assume that these males are neither orange, black nor tortoiseshell. He also states that tortoiseshell or black females mated to tortoiseshell males do not get tortoiseshell male offspring as one would expect by his hypothesis. An- other criticism of much greater weight than any of the foregoing is that he has Crossing over of a sex-linked factor taking place in the male, which is assumed to be heterozygous for sex. This is contrary to all known facts, for in those cases even in which a Y chromosome is known to be present it has never been demonstrated that it carries any hereditary factors.[2]
[2: Johannsen (1913, pp. 609-612) believes Doncaster is not justified in postulating sex chromosomes as bearers of the factors for black and orange. He therefore makes some modifications of Doncaster’s interpretation in order that the factorial treatment may be more "purely Mendelian.” Male cats are represented factorially as Mm and females as mm, and the factors for black and orange are assumed to be rather closely linked to m. Even with these modifications the hypothesis is essentially the samt as Doncaster’s and is open to practically the same objections.]
Whiting’s (1915) explanation for the unexpected color types is entirely different from Doncaster’s. He thinks that the black females in matings No. 1 and 3 (table 1) are in reality tortoiseshells which have had the black pigmentation extended to such a degree that little or no orange is visible. It is quite possible that some of the unexpected blacks were obtained in this manner. I have obtained a few "self” black guinea-pigs from tortoise X tortoise matings (Ibsen 1916), but it seems improbable that over 20 percent of the total female offspring in mating No. 1 should be accidental blacks, as would be necessarily the case on this hypothesis.
Whiting also discusses the possibility of white spotting affecting the phenotypic appearance of an animal. In a tortoiseshell the white areas may fall, he suggests, on those spots which would otherwise have been orange and thus leave only the black pigmentation visible. Such an animal would be a black-and-white in appearance. This condition is well known in guinea-pigs. Unless, however, the amount of white spotting is relatively large with respect to the amount of yellow the chance of its all being covered, so as to produce a black-and-white, is very small (Ibsen 1916). According to Barton (1908) a piebald (black-and-white) is usually white only on the face, breast and feet. If any of the blacks in mating No. 1, therefore, had been white even to this extent, it is probable that they would have been classified as piebalds rather than as blacks. Even though they were not so classified, it is still improbable that the usual small amount of white woüld have covered all of the yellow.
The rare tortoiseshell male, Whiting suggests, is "genetically a yellow with an extreme of black extension factors or a black with an extreme of yellow extension factors.” This hypothesis lacks definiteness, and is practically impossible to prove or disprove.
From what has preceded it is evident that the hypotheses offered are far from perfect in that they either fail to explain adequately all the facts or are difficult to test experimentally.
I have been attracted to the problem of inheritance in the tortoiseshell cat because of its resemblance to the tortoise guinea-pigs with which I have been working. In guinea-pigs the self black condition is dominant to tortoise, but it is evident that this is not the case in cats. Tortoise in guinea-pigs is due to a definite single factor, while in cats Doncaster and Little assume it is due to the interaction of the black and orange factors. Whiting postulates extension factors governing the relative amounts of black and orange.
It seems possible to explain many of the apparent anomalies of color inheritance in cats by assuming that the tortoiseshell coat is due to one definite factor, which I have called T, and which can act only in the presence of black (B), causing the black to be restricted to spots and leaving orange areas between. Two other assumptions are necessary in order to explain all the facts; first that black (B) is dominant to orange (b), as in guinea-pigs, rats, dogs, etc., and second, that under ordinary conditions T (tortoiseshell) is closely linked to b (orange). These points can be brought more clearly to mind by an inspection of figure 1, in which the factors are represented as being lineally placed on the sex chromosome. It will be noted that two sets of allelomorphs are assumed rather than one as was postulated by Doncaster and Little. It must also be kept in mind that since T acts only in the presence of B (black) it will have no visible effect on males carrying the b (orange) factor or on females homozygous for this factor.
So long as T and b remain on the same chromosome the hypothesis here presented is as efficient as Doncaster’s or Little’s in explaining the normally expected classes in the different matings. It is by means of the occasional Crossing over of the factors that the attempt will be made to explain the unexpected classes.
Figure 1. Diagrammatic representation of sex chromosomes bearing color factors showing normal condition in which T and b (orange) are postulated to be closely linked.
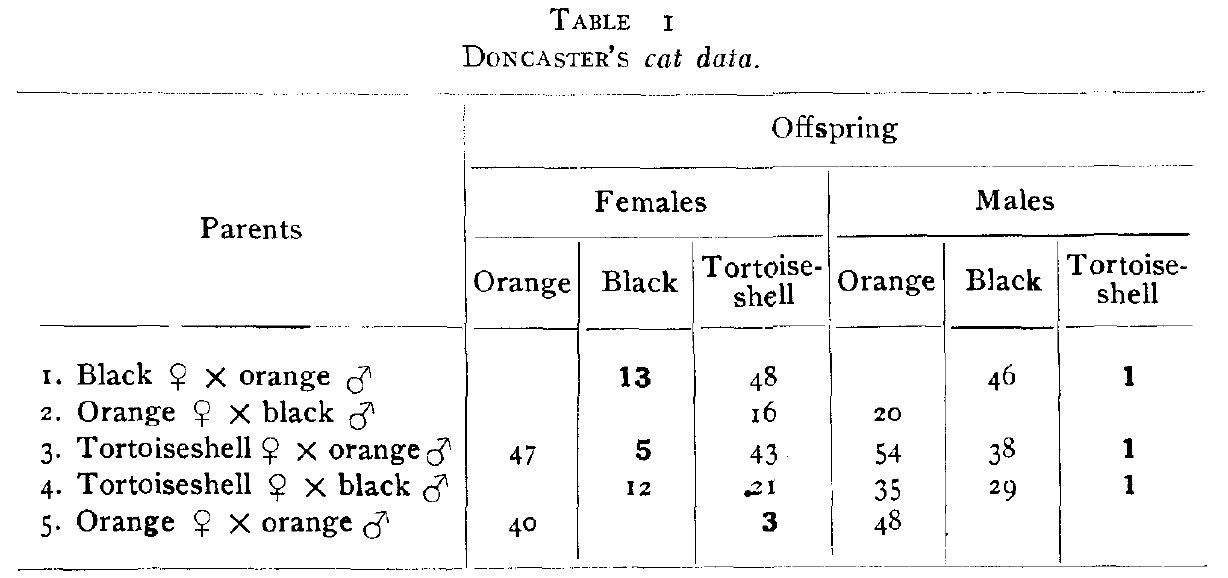
At the outset it may be stated that the hypothesis accounts for the unexpected black females and all the tortoiseshell males, except the one in mating No. 1, which, however, may be explained by a combination of Whiting’s hypothesis and my own. The tortoiseshell females in mating No. 5 are explained on Whiting’s hypothesis, which seems adequate in this particular case, as will be explained later.
I shall first take up the way in which tortoiseshell males may be produced. Since T and b are by hypothesis closely linked, we must assume that Crossing over takes place only rarely, and it can occur only in the female. Its occurrence in a homozygous female (figure 1, A or B) would produce no effect in the offspring. The heterozygous female (figure 1, C) is a tortoiseshell and normally forms gametes Bt and bT. When Crossing over takes place gametes BT and bt are produced. If this female were mated to an orange male, bT."—", the following classes would occur as a result of the Crossing over:
BT.bT, tortoiseshell female,
bt.bT, orange female,
BT."—", tortoiseshell male,
bt."—", orange male.
This would account for the tortoiseshell male in mating No. 3. With a black male, Bt."—"(figure 1, D), the offspring resulting from the Crossing over are,
BT.Bt, tortoiseshell female,
bt.Bt, black female,
BT."—", tortoiseshell male,
bt."—", orange male,
thus accounting for the tortoiseshell male in mating No. 4. Doncaster (1913) states that he knew of several tortoiseshell males besides those listed in table 1, all of which came from tortoiseshell females by unknown sires. It would therefore seem that tortoiseshell males, when they occur, come almost invariably from tortoiseshell females. This agrees with my hypothesis, according to which tortoiseshell males can be produced only by tortoiseshell mothers. The one recorded in mating No. 1 as having a black mother is an apparent exception. I have no explanation for this case, unless perhaps as already intimated the female was in reality a tortoiseshell, but with so little orange showing that she was recorded as a black.
In the paragraphs just preceding I have tried to indicate how tortoiseshell males are produced. I shall next show what kind of offspring are expected from them.
Doncaster records the mating of a tortoiseshell male with a black female. The female was not kept in confinement after copulation was observed, and Doncaster says there may be some doubt as to the paternity of the offspring. The only offspring recorded are a black male and tortoiseshell female, which are exactly what would be expected by hypothesis:"
Black female gametes: Bt bt
Tortoiseshell male gametes: BT "-"
F1: Bt.BT, tortoiseshell female
F1: Bt."—", black male
When tortoiseshell females are mated to tortoiseshell males Doncaster states that the following four types of offspring are obtained: tortoiseshell females, orange females, orange males, and black males. On my hypothesis no orange females should result.
Tortoiseshell female gametes: Bt bT
Tortoiseshell male gametes: BT "-"
F1: Bt.BT, tortoiseshell female
F1: bt.BT, tortoiseshell female
F1: Bt."—", black male
F1: bt."—", orange male
It will be noted, however, that the second type of tortoiseshell female in the F1 generation has a double dose of T and a single dose of B. It may be possible that the extra T restricts the black to such an extent that the animal appears to be an orange instead of a tortoiseshell. This is offered only as a suggestion.
A further test of the hypothesis would be to determine what kind of offspring are obtained from the F1 females resulting from the tortoiseshell x tortoiseshell cross. By hypothesis one-half of their male offspring should be tortoiseshells no matter what the color of the male is to which they are mated. There appears to be no record of a mating of this type.
We may next consider how the black females in matings No. 1 and 3 may have been formed. It will be remembered, that when crossing over takes place in the tortoiseshell female, the two kinds of crossover gametes formed are BT and bt, and that no matter whether the male parent is black or orange the male off spring are BT."-", tortoiseshell, and bt."-", orange. The bt."-" orange male is different from other orange males in that b is not linked with the T factor. By the mating of this sort of orange male with (1) a black or (2) a tortoiseshell female we should obtain black females which are otherwise unexpected in matings No. 1 and 3.
Black female gametes: Bt bt
Orange male gametes: bt "-"
F1: Bt.bt, black female
F1: Bt."—", black male
Tortoiseshell female gametes: Bt bT
Orange male gametes: bt "-"
F1: Bt.bt, black female
F1: bT.bt, orange female
F1: Bt."—", black male
F1: bT."—", orange male
It is to be regretted that Doncaster does not give the offspring from individual matings. If this had been done it would have been possible to test more thoroughly that part of the hypothesis dealing with the unexpected black females. It will be noted that when black females are mated to bt."—" orange males all the offspring should be black. There is no way of determining from Doncaster’s data whether or not this ever happens. Moreover, when tortoiseshell females are mated to bt."—" orange males none of the female offspring should be tortoiseshells.
Part of Doncaster’s data were obtained from Bonhote. These are now given separately in Bonhote’s (1915) recent book. Here the individual matings are presented and it is possible to trace the offspring of three orange males. Two of the males had the usual tortoiseshell and orange daughters when mated to either black or tortoiseshell females, while the third had 1 orange, 3 black, 6 blue and 7 tortoiseshell female offspring when similarly mated. This last mating does not fit in at all with the theory that the orange male parent was bt."—". It should be pointed out, however, that, as Bonhote states, he always selected tor-toiseshell mothers carrying a large amount of black, which would facilitate the production of tortoiseshell daughters with large amounts of black also. Some of these daughters might therefore have the appearance of blacks. It will also be noted that many (6) of the blacks were dilute and hence classified as blues. I have noticed in dilute tortoise guinea- pigs that it is much more difficult to detect the small yellow (dilute red or orange) spots surrounded by dilute black hair, than it is to detect the small red spots surrounded by deep black hair. It is therefore possible that in this way also animals which should have been classified as tortoises have been called blacks (blues).
The crossover (bt."—") orange male should be as rare as the tortoiseshell male. The latter is often sterile and it is possible the bt."—" orange male is sometimes sterile also. Taking all this into consideration it is probable that matings between either black or tortoiseshell females and bt."—" orange males are comparatively infrequent; thus it is quite possible that Doncaster has no record of this type of mating. If such matings occur, however, the black female off spring would be of the formula Bt.bt. These bred to either orange or black males should have orange and black sons in equal proportions. The orange sons would be bt."—". On Whiting’s hypothesis the unexpected black females should have orange and black sons also, but the orange sons should be bT."—". Doncaster states that he knows of "no satisfactory record of a yellow male mated to a black female having yellow sons.” From this one may infer that there have been cases reported in which a black female had orange sons, but none of them so far have been thought reliable. A larger number of records may furnish some reliable cases.
I have tried so far to account for the tortoiseshell males and the unexpected black males. The three tortoiseshell females in mating No. 5 remain to be explained.
Barton (1908), writing for the fancier, states, "If there is no white [in a tortoiseshell], then the amount of black hair should be small, compared with the red [orange] and yellow [dilute orange] markings.” According to Doncaster the three tortoiseshell females in mating No. 5 come from one mating. It is therefore possible that in selecting for a tortoiseshell female with the requisite small amount of black spotting one was obtained showing so little black that it appeared to be an orange. Mated to a true orange male this should produce some tortoiseshell female offspring. Whiting has given essentially the same explanation in his 1915 paper.
While it must be admitted that the foregoing hypothesis is not entirely satisfactory, and carefully controlled experiments are necessary for its substantiation, it still has two decided advantages, (1) it is quite definite, thereby tending to be comparatively easy to prove or disprove, and (2) it violates none of the accepted tenets of genetics.
GENERAL COMPARISON OF TRICOLOR IN GUINEA-PIGS, BASSET HOUNDS AND CATS
In parts I, II and III of the present series characteristics of the tricolor coats of guinea-pigs, Basset hounds and tortoiseshell cats have been considered separately with little reference to their resemblances or differences. They will now be considered briefly from this general comparative viewpoint. We shall first take up the spotting characters themselves, after which their factorial relations will be discussed.
The tricolor coats of guinea-pigs, and tortoiseshell cats show a much greater resemblance to each other than do either of them to the tricolor coat of Basset hounds. They will therefore first be compared with each other and then both can be compared with the Basset hound coat.
Since white spotting tends to mask the true black-red relationship, it is better first to consider black and red alone in their relation to each other, and then to take up white spotting as it affects both of them. In both cats and guinea-pigs the black spotting is variable in amount and is quite irregularly distributed. In both, black may be so far extended that the animal looks like a self black or so little extended that it resembles a self red. Where the animals differ from each other is in the white spotting. In tortoiseshell cats, as bred by the fanciers, this is small in amount and has therefore little chance of blotting out all of the orange (red) or all of the black. In guinea-pigs, on the other hand white varies greatly in amount and distribution. It can therefore at times blot out either the black or the red and in this way produce red-and-whites or black-and-whites instead of the usual tricolors. Thus we see in guinea- pigs that the variability in amount and distribution of both the black and the white spotting help to produce the unusual color types, while in cats this is affected only by the black spotting. Because of this, cats tend to have fewer of the unusual types than do guinea-pigs.
Basset hounds differ decidedly in several respects from both tortoiseshell cats and tricolor guinea-pigs in that black is localized on the back, so that if there were no white present the entire head, legs and belly would presumably be tan in color, making the animal a black-and-tan. There is no chance here for the black to be so far extended that the animal appears to be a self black or so little extended that it appears to be a self red. The distribution of the white is also somewhat different. In both guinea-pigs and dogs, as well as in mammals in general, pigmen- tation tends to recede toward definite centers, and each of these centers may become entirely devoid of pigmentation. The order in which the centers become pigmentless seems to be quite irregulär in guinea-pigs, while in dogs there is greater regularity. The point that concerns us in the present instance is that the aural patches are the last from which pigmentation entirely recedes, and as already noted, these patches in the Basset are always tan. As a consequence the dog may be entirely white except for these tan patches covering the ears. Such an animal is a tan-and-white. Black-and-whites never occur since black pigmentation is never found on the head in Bassets.
Summarizing, we find that in cats white spotting plays a very unimportant role in the production of red-and-whites and black-and-whites; black spotting is perhaps occasionally responsible for these color types. In guinea-pigs black spotting and white spotting are co-equal in their effects, while in Basset hounds white spotting is chiefly instrumental in the production of tan-and-whites, the black spotting merely being passive and aiding only by its position.
So far we have been considering in the three species only the visible relations of black and of white spotting to red. We may next take up their factorial bases. The white spotting factors in the three animals may have some resemblances so far as we know, but since white spotting has not as yet been adequately treated in a factorial manner, we are not in a position to discuss them. The black spotting factor, on the other hand, can be more definitely discussed.
In guinea-pigs this is called the partial-extension factor (ep) and is the middle term of a triple allelomorphic series of which entire extension (E) and non-extension (e) are the two extremes. It is therefore recessive to entire extension. In cats on the other hand the factor for black spotting is assumed to be a dominant partial-restricting factor (T), sex- linked, and also closely linked to the orange factor (b). Here it is dominant to entire extension of black. In Basset hounds two factors have to be considered. E is present just as in guinea-pigs, but black spotting here is not due to a modification of E as it is in guinea-pigs. Instead we have a new factor (T) which is not found in either guinea- pigs or cats. The T in cats and the T in dogs are two entirely different factors. As found in dogs it is the factor for the self-colored condition. In its absence (t) the animal is bi-colored. Since a bi-colored animal may be a red-and-lemon as well as a black-and-tan we see that t differs from ep in guinea-pigs and T in cats in that it has to do with spotting with red as well as spotting with black or chocolate. In order to get dogs spotted with black but not with red, E and T must both be present.
We thus see that though characters in different animals may show some resemblances, they may differ entirely in their factorial analysis.
LITERATURE CITED
[Messybeast Footenote: Doncaster should have been aware of the following fertile tortoiseshell male show cats: 'Champion Ballochmyle Samson,' owned by Lady Alexander, and 'Champion King Saul,' owned by Mrs Herring.

BALLOCHMYLE SAMSON - Short-haired tortoiseshell male. Born May, 1894
Dam: Mr. Shaw's Topsy. Sire not recorded. Parents’ colours not recorded.
BALLOCHMYLE MERMAID - Short-haired tortoiseshell-and-white female. Born March, 1897
BALLOCHMYLE BOUNTIFUL BERTIE - Short-haired tortoiseshell female. Born March, 1897
Sire: Ballochmyle Samson (tortie male). Dam: Mrs. Heslop's Topsy (colour not recorded)
Topsy’s colour is unrecorded so we can’t tell if Samson bred as an orange male or a black male.
KING SAUL - Short-haired tortoiseshell male
Sire: Darkie. Dam: Flossie (common names, cannot be traced further back) (colours not recorded)
KING ORION - Short-haired orange male. Born May, 1899
NORA - Short-haired black-and-brown (tabby?) female. Born July, 1900
Sire, King Saul (tortie male). Dam. Lady Jasmine (short-hair tortie)
By modern analysis, King Saul bred as a black male.]
INHERITANCE OF COAT COLOUR IN CATS - Phineas W. Whiting 1st August 1918
CONTENTS
I. Introduction: The color-factors of domestic cats
II. Presentation and discussion of data
A. Maltese dilution
B. White and white-spotting
C. Solid yellow and yellow-spotting
D. Siamese dilution
E. Banding and ticking
E-a. Statement of factorial differences and description of characters
E-b. Experimental data
E-c The number of loci involved
E-d Physiology of color-production
III. The origin of color varieties of the cat
IV. Summary
V. Literature cited
INTRODUCTION: THE COLOR FACTORS OF DOMESTIC CATS
In a series of experiments begun at the University of Pennsylvania in the autumn of 1914 and extending up to the present time, the inheritance of coat-color in cats has been investigated. Although the number of litters obtained has not been large, it has been found possible to determine several points in regard to the mechanism of heredity by means of critical crosses. This has been largely due to the fact that the characters studied segregate for the most part cleanly from each other so that it has been easy to classify the animals.
My thanks are due to Dr. McClung and to Dr. Colton for the kindly interest which they have taken in the work and to the University of Pennsylvania for the expense of the experiments. I also wish to thank the Zoological Society of Philadelphia for the opportunity of crossing my cats with the Caffre cat.
Before presenting the data and discussing the inheritance of the various characters in detail it may be well to name and to define briefly the factorial differences involved.
PRESENTATION AND DISCUSSION OF DATA
A. Maltese dilution
Maltese dilution appears to be a simple Mendelian recessive. It apparently exists in combination with all other factorial differences, but I have not as yet seen its representative in the lined or narrow type of banding. It is always sharply distinguishable from black, but shows considerable variation in its own intensity. It is to be compared to slaty-blue in the mouse, the rabbit, and the dog. No corresponding color is known in the rat or in the guinea-pig. Cream or dull yellow is its corresponding color in the yellow series; blue and cream, in the tortoiseshell.
These data show merely that maltese dilution is not sex-linked.
B. White and white-spotting
White-spotting in cats is exceedingly irregular in amount and distribution, but tends to appear more commonly on the under parts. There appears to be no regularity in dominance and probably many factors are involved. The degree of white- spotting in the parents tends to appear again in the offspring, although wide segregation occurs. I have usually selected cats with relatively small amount of white and have obtained kittens of the same general character.
Solid white appears to be a complete dominant over color whether the color is self or spotted with white. It is possible that it may be allelomorphic with one or more of the white- spotting factors, but my data are not conclusive on this point. Castle (1916) regards it as possibly an extreme form of white- spotting.
A cream male (8)[Individual numbers have been inserted after mention of any animal that is referred to more than once in this paper. Matings have been numbered similarly. It will thus be possible for the reader to check up the genetic composition of any animal by its progenies from various matings] with slight amount of white crossed to a blue-eyed deaf white female (7) sired (6) three pure white kittens; one male and two females. The kittens had normal hearing and developed yellow eyes. The male had a slight smuttiness of the hairs on top of the head which appeared when he was two weeks old and then shortly disappeared. The females never developed any pigment in the hair. The same pair of cats mated again and produced two solid white kittens which were not reared.
The cream male (8) was later mated (32) to a yellow-eyed white with normal hearing (22). Two whites, a male and a female, were produced, as well as two females that were entirely self color, a blue and cream and a cream. I was unable to detect any white hairs on these two pigmented kittens. It would appear, then, that the white mother carried factors which dominated the slight white marking of the father, and thus produced totally self-colored kittens.
Dr. Little reports a mating (43) of a yellow-eyed white male by a cream and white female. A single yellow-eyed white female was produced.
A mating (35) of a blue-eyed white Angora male (16) by a yellow-eyed short-haired white female produced three solid whites and a black that was self-colored except for very small white spots between the legs. A mating (36) of the same male by a maltese female produced a solid white and a near self black.
Besides these I have three records of blue-eyed white females which produced both colored and white offspring. The male parents were unknown. One produced a solid white yellow-eyed female and a solid black male. One produced in two litters six solid white and three almost entirely black. The third produced a solid white and an intense striped tabby with belly, nose, breast, and feet white.
Davenport (1905) reports a mating of a black Manx male by a blue-eyed, deaf, white female, which produced two whites, one black, one tiger, and one maltese.
The interesting correlation of blue eyes and deafness with white coat is not yet satisfactorily explained. Dominance of eye color seems very irregular. I am informed by breeders of white cats that yellow-eyed by yellow-eyed may produce blue-eyed, and also that blue by blue may produce yellow. Odd-eyed cats also frequently appear in these crosses.
Przibram (1908) reports experiments with odd-eyed white Angora cats. Results were very irregular. The cats bred true to whiteness. As regards correlation of deafness and blue eyes, he says, "It is interesting that, as far as a hearing test could be undertaken, the blue-blue adult cats were all deaf, which is consistent with earlier observations by Darwin and Rawitz; the asymmetrical eye-colors seem to be affected with a one-sided deafness affecting the side with the blue eye. The correlation between blue eyes and deafness thus also persists in asymmetric inheritance. The correlation between 'blue eyes' in cats and 'deafness' also persists for the body halves.”
[Von Interesse ist es, dass, soweit eine Prüfung des Gehörs unternommen werden konnte, die blau-blauen erwachsenen Katzen alle vollig taub waren, was ja mit früheren Beobaehtungen von Darwin und Rawitz übereinstimmt; die asymmetrischen Augenfarben scheinen dem ganz entspreehend mit einer halbseitigen, die Seite des blauen Auges betreffenden Tau'bheit betroffen zu sein. Die Correlation zwischen blauen Augen und Taubheit bleibt also auch bei der asymmetrischen Vererbung bestehen. Dabei bleibt die Correlation zwischen ‘blauen Augen’ der Katzen und ‘Taubheit’ auch für die Körperhälfe bestehen.]
Dr. C. C. Little showed me a black and white cat with odd eyes. The hair surrounding the blue eye was white, while that about the yellow eye was black. Blue eyes in pigmented cats are rare, except of course in the case of the Siamese.
I would suggest, therefore, as a working hypothesis that the incidence of white-spotting in connection with the dominant white factor produces the blue eye, or in other words a 'white spot’ about the eye of a white cat makes the eye blue, while a ‘pigmented spot’ about the eye of a solid white cat makes the eye yellow. It may be also that a ‘white spot’ in the ear of a white cat makes it deaf. This would explain why it is so difficult to get blue-eyed white cats with normal hearing as it would be difficult to localize the ‘white spot' upon the eye and to keep it away from the ear. This may also explain why odd-eyed cats are frequently defective in hearing only on the side having the blue eye, as noted by Przibram. It would not be a difficult matter to test this hypothesis.
C. Solid yellow and yellow-spotting
The tortoiseshell cat has been the subject of much interest and discussion in genetic literature dealing with sex-correlated phenomena. Doncaster (1905) considered the problem and tried to explain the peculiarities of inheritance by variations in dominance. Little (1912) suggested the hypothesis of a single sex-linked pair of allelomorphs with the male digametic. He used the term ‘sex-limited character,’ which has since been restricted to simple Mendelian heredity in which sex reverses the dominance of the allelomorphs. Doncaster (1912) accepted Little’s suggestion as in general satisfactory, but pointed out that occasionally black females are produced from matings of black female by yellow male. According to Little’s hypothesis, the females should always be tortoiseshell from the reciprocals of black by yellow and the males should be like the mother, disregarding of course dilution, tabby, etc. Doncaster suggests an occasional break in sex-linkage to explain these anomalous blacks, as also the occurrence of the rare tortoiseshell male. In 1913 he gave a further discussion of the subject and an excellent summary of data collected from fancy breeders. In 1914 he suggested nondisjunction of the sex-chromosomes in oogenesis to explain the matroclinous black females. These explanations are all more or less unsatisfactory for one reason or another, as admitted by Doncaster and by Little.
I have pointed out (1915) that the hypothesis of simple sex- linkage first suggested by Little may be sufficient to account for the conditions if it be considered that the heterozygotes, which must be females, vary from black through various degrees of yellow-spotting to solid yellow. In the male, presumably, conditions are much more stable, as it is impossible to have a heterozygote. Thus I have suggested that a gametically yellow male (YX —) may become tortoiseshell by extreme selection of black extension factors, while a gametically black male (yX —) may become tortoiseshell by an extreme selection of yellow extension factors. The possibility is of course not excluded that there may be a single factor or particular combination of factors that produces yellow-spotting in the male.
Ibsen (1916) has suggested close coupling of two pairs of sex- linked allelomorphs, and attempts to explain anomalies by crossing-over. This does not account for all the results, however, as he himself points out.
The data concerning the tortoiseshell problem which I have gained from my experiments are as follows:
Summarizing matings of 'yellow’ male (YX —) by ‘black’ female (yX yX), we have seven matings giving fifteen ‘black’ males (yX —) and thirteen ‘ tortoiseshell’ females (YX yX).
Crosses of yellow male by tortoiseshell female are as follows:
Summarizing crosses of ‘yellow’ males (YX —) by ‘tortoiseshell’ females (YX yX), we have four matings giving three ‘black’ males (yX —), three ‘yellow’ males (YX —), four ‘yellow’ females (YX YX), and four ‘tortoiseshell’ females (YXyX).
The long-haired cream male (8) (a.b.m.Y) mentioned above was crossed (32) to a yellow-eyed white cat (22). There were produced one white male, one white female, one cream female (B.m.YY), and one blue and cream female (a.B.m.Yy). Since a cream kitten as well as a blue and cream was produced, it is probable that the yellow-eyed white was gametically a tortoiseshell.
Dr. C. C. Little has very kindly supplied me with data in regard to an anomalous cream female (23) which breeds like a tortoiseshell. This female he has given to me along with three of its offspring. She has, while in my possession, produced four kittens by her cream son. They are two cream females, one cream male, and one maltese male. The maltese male would of course not be expected from a mating of two yellows. The under parts of the anomalous female are white. The upper parts are entirely cream and show the blotched pattern very plainly. No trace of black pigment can be found, although I have examined samples of the hair from various parts of the body under the microscope.
The following gives in detail the offspring from this anomalous cream female. The unexpected progeny are recorded in italics.
Summarizing the matings of this cream and white female, we find that:
Of these matings Dr. Little says: "The dilute yellow and white female is interesting because she forms gametes carrying black and breeds exactly like a dilute tortoiseshell and white animal, although there is no trace of black pigment anywhere on her.” She is then an anomalous yellow. Dr. Little further states: "Dilute yellow, like the same color in mice, does not depend upon the depth of color, but is essentially a dull yellow ranging anywhere from intense pigmentation to dilute cream color.” It is of course relatively not as intense as the orange. It is possible that this variation in cream color is due to the same factors which produce the variations towards silver in tabbies and others.
Yellow-spotting in cats may be compared essentially to the same condition in guinea-pigs. In the latter there is great range of variability as in the former. In cats, however, one of the allelomorphic pairs determining black or yellow extension is much more potent than the others and is sex-linked. The heterozygous female (YX yX) represents a much more unstable condition as regards spotting than either of the homozygous females or than either of the haploid males, for in the heterozygote the factors yellow, Y, and black, y, are balanced against each other.
The sterility of the tortoiseshell tom has frequently been remarked upon. Cutler and Doncaster (’IS) discuss this question and show drawings of sections of the testis of a sterile cat of this sort. Normal reproductive cells are altogether lacking. In summarizing the data on sterility of male tortoiseshells, they find that one was certainly fertile, two completely sterile, one almost if not quite sterile, and two doubtful. It appears, then, that sterility may be highly correlated with yellow-spotting in the male.
The black-yellow allelomorphic pair in cats is of particular interest, as it is the only case of sex-linkage known in mammals, other than the sex-linked defects of man.
D. Siamese dilution
Bateson (1913) says of the Siamese cats: "These animals, which breed perfectly true, were introduced from Siam, where they have been kept for an indefinite period as pets of the royal household. Like the Himalayan rabbit, Siamese cats are born almost white, but the fur becomes a curious fawn with darker chocolate points on the ears and extremities.” Crosses of Siamese by other cats are cited by Weir (1889). Quoting from a Mr. Young, he says (p. 76), "They invariably showed the Siamese cross in the ground color.” But Lady Dorothy Nevill says, "None showed any trace of the Siamese, being all tabby.”
Two pregnant females of common cats brought into the laboratory produced kittens of a peculiar ashy color with darker extremities. The kittens resembled very closely adult Siamese cats.
One of the pregnant females, a maltese (5) (a.m.) produced (15) two females which were ashy, with nose, ears, feet, and tail slightly darker, and two females and two males which were ashy with black extremities. A record taken fifty days after birth showed that the lighter kittens had become maltese, while the kittens with black extremities had become steel colored or almost black. They later became completely black. Ghost patterns were seen on four of the kittens, but unfortunately a critical examination was not made of the other two. Ghost blotched was very much accentuated by the ashiness, but ghost striped did not appear especially so. It is probable for this reason that the two doubtful ones were ghost striped. Of the other four the maltese was ghost striped, and the three steels were ghost blotched.
The other pregnant female (2) was a black and white (a.M). She produced (4) four kittens—two black females upon which no record of ghost pattern was made, an ashy female with black extremities, and an ashy male with dark but not black extremities. Sixty-six days after birth the ashy female had developed into a steel black which clearly showed ghost-blotched pattern, and eighty days after birth the ashy male was maltese with ghost-blotched pattern very evident.
Unfortunately, the inheritance of this peculiar ashy color could not be followed out at the time the kittens were on hand. I am, therefore, unable to say whether it represents the heterozygote for Siamese dilution.
E. Banding and ticking
a. Statement of factorial differences and description of characters.
Ticking or agouti in cats, as in rodents, is characterized by yellow bands on the hairs. It increases with age so that kittens are relatively less ticked than cats. I have tentatively considered the agouti factors as a series of triple allelomorphs — A', much ticked, A, little ticked, and a, non-ticked, with dominance of A' over A and a, and of A over a.
The banding factors, I have also represented as triple allelomorphs — B', lined, B, striped, and b, blotched. These factors affect the formation of yellow pigment, in a yellow cat (A or a.B.M.Y) forming bands of straw color alternating with orange. In a tortoiseshell tabby cat (A.B.M.Yy) the orange bands in the ‘yellow spots’ are continuous with the black bands in the ‘black spots,’ while the straw-colored bands are Continuous through both regions. In a tortoiseshell (a.B.M.Yy) alternate banding of straw and orange shows clearly in the ‘yellow spots,’ while the ‘black spots’ are uniform black. The same condition obtains in the case of maltese dilution, but the contrast in the bands is not as obvious and there is general reduction in the amount of yellow pigment.
Uniformity or lack of banding in yellow cats is apparently due, as has been pointed out to me by Dr. Sewall Wright, to some other condition than the lack of the agouti factor. As regards the existence of such cats, Mrs. Leslie Williams (1908) writes: "The self-orange Persian is more of an ideal than a reality, for it is actually a red tabby without the tabby markings, and at present it is a case of ‘more or less,’ the upshot being that the least marked cat in the class takes the prize.”
Silvering is a general reduction in the amount of yellow pigment. The straw bands of tabbies then become white. Figure A shows a silver-striped tabby skin. Black stripes alternate with white. In the skin shown in figure B, on the other hand, there is a considerable amount of yellow pigment. A striped tabby (9) brought into the laboratory pregnant had lighter bands of a decidedly reddish color. This apparently represents the opposite extreme of variation from silvering. Intense black stripes alternated with rusty red. She gave birth (22) to three male kittens—one striped with black and red; one blotched with black and red, and one striped with black and straw color. Here, then, is an indication that the extreme reddish tone is hereditary.
For an understanding of banding we may first consider figure B. The skin shown here is from a striped tabby male (7.2) forty-two days old. It may be seen that the bands run longitudinally along the back and are most easily seen near the mid-dorsal line posteriorly. On the sides the bands are transverse and tend to be broken into spots. We may think of this condition as having been produced by longitudinal and transverse waves of pigment-forming metabolic activity. The longitudinal waves form transverse bands. The areas of greatest activity form orange bands in yellow cats, while in tabbies these bands are black. The areas of less activity form, of course, the lighter bands. The transverse waves appear to originate at the mid-dorsal line. They form longitudinal bands on the back. As they pass outward and down the sides, the areas of greater activity tend to thicken the transverse bands. In the areas of less activity the transverse bands are often evanescent. It thus appears that black or orange spots, in tabbies or yellows, respec-tively, are produced in the regions of greatest metabolic activity.
The ticking and the banding factors appear to act in the same regions, and thus the ticking reveals the straw-colored rather than the orange bands. Agouti is, however, in all probability uniform over the body surface in cats as in rodents. This matter will be discussed in detail after the presentation of data.
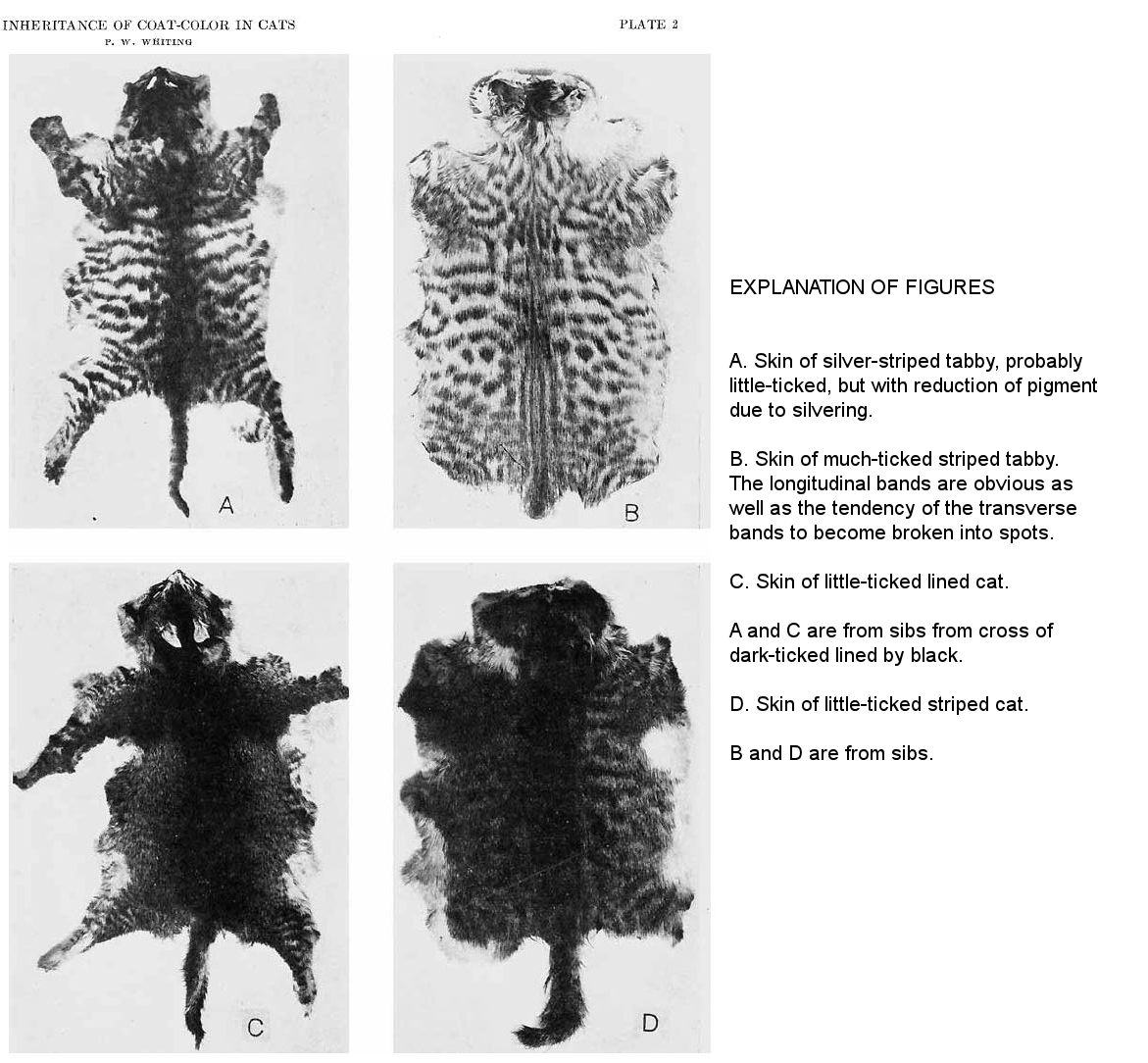
The skin shown in figure B had a high degree of ticking, and thus shows the longitudinal bands clearly. The cat shown in figure A is less ticked and the increased amount of black pigment on the back obscures the longitudinal bands. Figure D shows a very dark-striped tabby. While the bands on the sides are clearly seen, the longitudinal bands are obliterated by the black pigment. The cats shown in figures B and D are from the same litter and represent extreme segregation of ticking.
As has been said, ticking increases in cats as in rodents with maturity. The same kitten may, therefore, show different degrees of it at different ages. It is thus necessary to consider age in making comparisons with respect to this character. Ticking always segregates sharply from black. Various degrees of ticking ranging from that shown in figure B to that shown in figure D, however, occur. I have classified the extremes tentatively as A' and A, but their allelomorphism with "a” is uncertain. There may be intermediate allelomorphs or the variations may be due to modifiers.
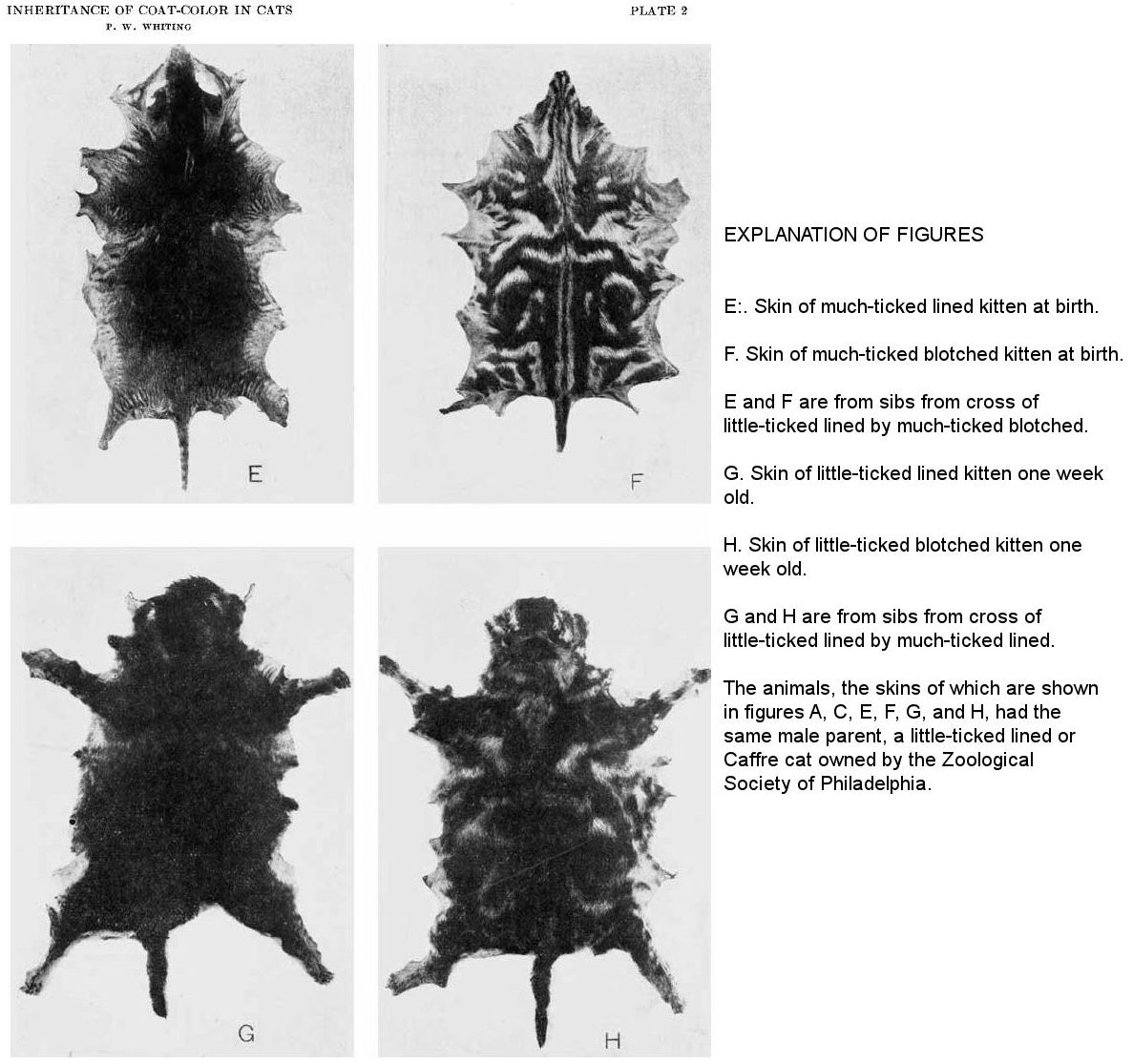
The blotched pattern is shown in figures F and H. Figure F is from a kitten extremely ticked at birth. Such a kitten develops into a cat that has yellow in all of its hairs. The black bands of the kitten become ticked in the adult. The lighter bands become entirely straw-colored. We have in this extreme ticking an approach toward the sooty yellow, as in the mouse.
The skin shown in figure H is from a kitten one week old. Nevertheless, it is much darker than that shown in figure F. Such a kitten develops into a dark-blotched adult. The ticking increases with age until the cat appears much like the kitten shown in figure F.
For a discussion of the blotched pattern in comparison with the striped, the degree of ticking shown in figure F is most favorable. The bands shown here are broad and consequently not as numerous as in the striped. A median dark longitudinal band down the back is cut just behind the shoulders by dark and light transverse bands. The alternation of dark and light bands is not as obvious in the blotched pattern as in the striped, since the bands are relatively wide and the longitudinal and transverse bands interfere with each other. The tendency of the bands to become broken into spots or blotches may be explained in the blotched, as in the striped, by a conflict of longitudinal and transverse waves.
The lined or narrow-banded pattern is rarely seen in cats in this country. The bands are extremely narrow and frequent and are best seen when the hair is very short and the ticking is of just the proper degree. Figure C shows a rather dark-lined cat (28.3) forty-five days old. The narrow banding shows clearly about the edge of the skin and to some extent on the sides. Such a cat becomes somewhat lighter when it grows older. It is very dark when young and appears black and tan like figure G, which is from a lined kitten (19.3) one week old. Narrow bands are seen in the tan areas of the latter and the back and sides show narrow bands when the skin is turned in certain relations to the line of vision. These narrow bands are really a ‘ghost pattern’ comparable to the ‘ghost patterns’ of striped and blotched seen in fully black cats. They may be seen in the fur running transversely down the sides. On the skin they may be seen running in the same way and also longitudinally down the back. They are much narrower and more numerous than the bands of striped cats. Lined cats occur in Africa and to some extent in Europe. They are known as African, Caffre, or Abyssinian cats.
In black and maltese kittens ‘ghost patterns’ are seen clearly in the skin and are not difficult to recognize in the fur. As the kittens become older the ghost patterns sometimes show more clearly in the fur for a time, although they disappear from the skin. In adult cats ghost patterns are occasionally seen, I have been able to classify all black or maltese kittens as either striped or blotched. A lined cat lacking agouti has not yet been obtained, but this I am hoping to do in time by the proper crosses.
Figure E is from the skin of a lined kitten at birth. It is an extremely ticked example and would probably have grown to a sooty yellow adult. The back is black, but well scattered with ticked hairs, thus differing from the skin shown in figure G. The transverse bands are shown about the edge of the skin at the sides and about the tail. The longitudinal bands are suggested by two ticked spots at the back of the neck. Just posterior to these spots are two parallel ticked lines. On the body near the tail may also be seen longitudinal bands.
Fundamentally, then, the lined, the striped, and the blotched patterns are comparable, differing only in the width of the bands.
A pair of lined cats is owned by the Zoological Society of Philadelphia. The male is dark while the female is much lighter. A comparison of the degree of ticking of the two may be of interest. The back of the male is black, the sides very dark showing narrow ticked bands. The back of the female is dark but ticked and grades into sooty yellow on the sides, showing no dark banding. The banding on the head and breast of the male is for the most part black, while in the female it is brown shading to sooty. In the male the back and end of the tail are black, while ticked rings are seen only toward the base. In the female the entire tail is ringed with sooty yellow. In both animals the feet are sooty yellow, the soles black. In the male the black bands of the sides extend down the legs to the feet, while in the female the sooty yellow of the feet extends well up on the legs.
b. Experimental data.
When bred together (19) the lined cats produced four kittens — a dark-lined male, the skin of which is shown in figure G; a dark-blotched male, the skin of which is shown in figure H; a dark-blotched female, and a light-lined male (21). The last-mentioned animal has been raised and is now in the possession of Dr. Charles Penrose, of Philadelphia. It has been examined by the writer, who finds that at the age of two years and three months it is in color almost the exact counterpart of its mother. It has, therefore, received from its mother the factor A' while its three sibs have received the factor A for which the mother is presumably heterozygous, unless indeed she is carrying non-agouti, a. The cross may be expressed: AAB'b x A'AB'b = 1 AAB'? + 1 A'AB'? + 2 AAbb. Whether or not the two lined offspring ate homozygous for B' or carry b is unknown.
The dark-ticked lined male was bred to a very much ticked blotched female (14). This female had been brought in preg-nant and had produced (11) two blacks showing ghost striped, one blotched, and two striped. One of the striped had a very high degree of ticking. The skins of the others were not saved and no determination of the exact amount of ticking was made at the time. When bred to the dark-ticked lined male this female produced four kittens — one lined and three blotched. The lined is shown in figure E and one of the blotched in figure F. The other two blotched were similar in amount of ticking to that shown in figure F. This female may then be considered as of formula A'abb. The cross to the lined male, AAB'b, gave 1 A'AB'b and 3 A'Abb. Larger numbers would probably have shown some dark-ticked kittens.
The same male was bred to the black and white female (2) mentioned under the discussion of Siamese dilution. She had produced (4) by an unknown male two ghost blotched kittens — a black and a maltese — and two blacks in regard to which no ghost pattern was recorded. When bred to the lined male she produced (29) four kittens—three lined and one blotched. All the skins were kept and all were very dark. The mating may therefore be represented: male AAB'b x female aabb = 3 AaB'b + 1 Aabb.
The lined male was also bred to a black female (10) which showed indistinct stripes in her fur. She had produced (23) from an unknown male four black males, all showing stripes as ghost pattern. When bred to the lined male she produced (28) one lined and two striped kittens. The lined is shown in figure C and one of the striped in figure A. The other striped was about the grade of ticking as that shown in figure A. If my hypothesis of the allelomorphism of the agouti factors is correct, these striped kittens must represent a light variation in the dark agouti. In the case of these kittens the light color may be due to silvering, as there is practically no yellow in the fur of either of the striped, although the lined shows a fair amount. The cross may be represented: male AAB'b x female aaBB = 1 AaB'B + 2 AaBb.
A highly ticked blotched male (11) was crossed (14) with a black female (15). Four highly ticked blotched kittens similar to figure F resulted. The cross may be represented: male A'A'bb x female 9 aabb = 4 A’abb. The same male was later crossed (25) with a maltese striped female (4). Three intense striped and two intense blotched resulted. Unfortunately, the grade of the ticking was not determined.
A very dark striped female (3) was crossed (16) to a very dark blotched male (6). Six kittens were produced—two blotched, three striped, and one black showing stripes as ghost pattern. The blotched and two of the striped were dark. The other striped was slightly lighter. It was not, however, as light as those that are grouped as much-ticked. It shows clearly, nevertheless, that ticked cats may produce offspring apparently more ticked than themselves. The cross may be represented: male Aabb x female AaBb = 3 AA or AaBb + 2 AA or Aabb + 1 aaBb.
The same dark-striped female (3) was crossed (33) with a longhaired cream male (8) which appeared from other crosses to be homozygous for blotched and for non-agouti. The two kittens were a blue and cream female showing blotched as ghost pattern and a dark-striped male. The latter has grown up and shows exactly the same dark coat-color as his mother. The cross may therefore be represented: male (aa.bb.mm.Y-) x female (Aa.Bb.Mm.yy) = 1 male (Aa.Bb.Mm.y-) + 1 female (aa.bb.mm.Yy).
The dark-striped female (3) was mated twice (34 and 49) with a short-haired cream blotched male (24) and produced nine kittens as follows: Males, 1 a.B.M.y., 2 a.B.m.y., 1 A'.B.M.y., 1 a.b.m.y. Females, 1 A'.B.m.Yy., 2 a.b.m.Yy., 1 A.b.M.Yy.
The light-ticked kittens probably inherited the factor A' from their yellow father. That the male carries the factor A', is shown by the fact that when crossed (45) with a black and orange (30) there were produced three males — a black and white in which there was so much white that it was impossible to determine the ghost pattern, a cream, showing stripes, and a blotched maltese in which the ticking stood out extremely clearly. The latter is being raised and differs decidedly in amount of ticking from many other blotched maltese cats in my possession.
The long-haired cream male (8) was crossed twice (9 and 30) with a black female (20). Ten kittens were produced, all of them showing stripes as ghost pattern. The female is therefore aaBB. The cross may be expressed: male aabb x female 9 aaBB = 10 aaBb.
The same male (8) was crossed (12) to a much ticked striped female (25). Of the four kittens produced two were non-ticked and showed stripes as ghost pattern and two were much ticked and striped. The cross may therefore be expressed: d1 aabb X 9 A'aBB = 2 aaBb + 2 A'aBb.
The same male was crossed (13) to a maltese female (17). Three dilute kittens resulted, all with blotched ghost pattern, c? aabb X 9 aabb = 3 aabb.
It is clear, then, that the cream male (8) does not carry the ticking factor, for when crossed with two non-ticked females it has sired thirteen non-ticked kittens. It also probably is homozygous for blotched, although it does not show the pattern, for when crossed to a maltese female it sired three blotched kittens.
It thus appears that the long-haired cream (8) does not carry agouti and does not show the blotched pattern, while the shorthaired cream (24) carries strong agouti, A', and shows the pattern. Whether the agouti factor tends to bring out the pattern more strongly in a cream may yet be an open question. I am inclined to think that the pattern is obscured to some extent in long-haired cats by the length of hair.
An orange-striped male was crossed (37) to a blue and cream female (26) that showed ghost blotched pattern. Two orange- striped females resulted. The cross may be expressed: male (BB.Y —) x female (bb.Yy) = 2 (Bb.YY).
c. The number of loci involved.
The facts thus far collected, then, are consistent with the assumption of two loci, one for the banding factors and one for the ticking factors. Since, however, all variations from very light to very dark ticking occur and since dark-ticked cats heterozygous for black may produce kittens of somewhat lighter grade than themselves, it is probable that factors at other loci recombine to modify the ticking. Tests are now being made which, it is believed, will determine definitely whether light ticking and dark ticking are both allelomorphic with the same factor for black; that is, whether they form a triple allelomorphic series.
The three types of banding, lined, striped, and blotched, are each entirely distinct. No intergrades have been observed. The natural assumption is to suppose that they form a triple allelomorphic series, B', B, and b, as I have tentatively assumed. But if two loci are involved the conditions might be expressed as follows: A lined cat might be LLBB, LLBb, LLbb, LlBB, LlBb, or Llbb. A striped cat might be llBB or llBb. A blotched cat would then be the double recessive, llbb. This scheme apparently fits the genetic data thus far collected. Striped and blotched would act as a pair of simple allelomorphs, B and b. The crosses involving the lined cats would be expressed by supposing that they are both of formula Llbb. Bred together they produced lined, LLbb or Llbb, and blotched, llbb. Crossed with blotched, either black or ticked, llbb, they give lined, Llbb, and blotched, llbb. Crossed with homoyzgous striped, llBB, they give lined, LIBb, and striped llBb.
The same crosses for testing the agouti factors will also test the allelomorphism of the banding factors. Other combinations that are being made with lined shouild give yellow and maltese lined.
d. Physiology of color-production.
Reference should now be made to Wright’s (1917) papers on color inheritance in mammals. Wright classifies color factors according to their effects on either one of two enzymes. Enzyme 1 is the basic enzyme for color production which, acting alone on chromogen produces yellow. Enzyme 2 is supplementary to enzyme 1. It has no effect alone either on chromogen or on yellow pigment. Combined with enzyme 1 it oxidizes chromogen to sepia.
The agouti factors are considered as determining an inhibitor of enzyme 2. "Factor A determines the production of an inhibitor with the same subtraction effect on enzyme 2 everywhere.” This inhibitor acts in waves along the individual hairs. The regions of greatest concentration determine the yellow bands, while those of less concentration are black.
In yellow cats it is seen that banding occurs over the surface of the body, straw-color alternating with orange. Banding, therefore, affects enzyme 1. In black eats the bands are almost indiscernible. There is, then, enough of enzymes 1 and 2 generally distributed to produce a uniform black. In the presence of the agouti factor, however, yellow bands appear in the individual hairs. These bands are much wider in the areas corresponding to the straw-colored bands of yellow cats. In fact, the black may be here entirely obliterated. The hairs in the areas corresponding to the orange bands of yellow cats are much darker and may be without apparent ticking. It therefore appears that the banding factors affect enzyme 2, for if enzyme 2 were uniformly distributed as in rodents, the agouti factor should cause a uniform ticking over the body surface, not an alternation of dark and light bands.
The banding factors may be thought of, then, as determining waves of general metabolic activity affecting both enzyme 1 and enzyme 2. In the black eat the regions corresponding to the orange bands in the yellow cat would be a dense black, a sort of black dominant to agouti, comparable to Punnett’s (1912 and 1915) dominant black in rabbits, while the regions corresponding to the straw-colored bands would be comparable to ordinary black in being recessive to agouti.
For helpful criticism and discussion of these matters I am much indebted to Dr. Wright, whose papers I have already mentioned.
III. THE ORIGIN OF COLOR VARIETIES OF THE CAT
It is generally assumed that the domestic cat is polyphyletic in origin. Darwin considered this to be the case. Keller (1902) discusses the matter and agrees with Darwin on this point. Elliot (1883) believes that the cat is descended from a number of wild species and supposes that it has crossed at various times with small wild cats in different countries. He attempts to trace the well-known color variations as well as variations in form to such hybridizing.
Rope (1881) and Pocock (1907) both recognize the characters blotched and striped and believe that all cats, whatever their color, fall in one or the other of these two classes. Pocock states:
It is needless to say more in support of the contention that if a decided difference in the ‘pattern’ of Domestic Cats exists, it must be regarded as furnishing a surer basis for their classification than the length of hair, the tint of the coat, or the stunting of the tail. It may also be claimed with assurance that the pattern supplies a more im-portant clue to the ancestry of the Domestic Cat than the features
just mentioned Frequently at all events the so-called
‘blotched’ pattern can be detected in certain lights even in 1 Whites’ and ‘Blacks.’ "
Pocock also recognized the lined variation, called by him Abyssinian. "Cats of the so-called ‘Abyssinian’ breed may be descended, for anything I know to the contrary, from specimens of F. ocreata directly exported from Abyssinia. They are certainly not unlike some self-coloured examples of that species. On the other hand, it would, I imagine, be difficult to separate them from fulvescent ‘Ticked’ Cats, which appear to me to be nothing but examples of the torquata-type in which the pattern is broken up and evanescent.”
The torquata type is what I have called striped. Pocock discusses the synonymy of wildcats and of the domestic cat. The whole matter appears to be much confused.
[Footnote 3. The wildcat of Europe is usually called Felis catus L., but inasmuch as Linnaeus’ description agrees with the blotched pattern while the European wildcat is striped, it is considered by Pocock and others that Linnaeus was referring to the domestic cat. Felis sylvestris Schreber is therefore chosen as the name for the European wildcat. Of the striped form Pocock says: "To feral or domesticated examples of this oat have been given many names, of which torquata is the best known and angorensis or striata possibly the oldest. It closely resembles in pattern two existing species, namely, the so-called Egyptian cat (F. ocreata) and the European wildcat (F. sylvestris).” Pocock thinks that the blotched or catus type is derived from some extinct, probably Pleistocene cat of western Europe. Pocock uses the term torquata for striped and catus for blotched, which is just the reverse of many other authors. There is, further, a so-called Felis torquata of India that is considered by some to be related to the spotted desert cat (F. ornata). Inasmuch as I have wished to name genetic factors rather than species of cats, I have discarded the Latin names and have adopted the English words blotched and striped, in regard to which there can be no confusion.]
Dr. A. Nehring (1887) believed that the cat has a dual origin, being descended from a domestic Chinese cat and from the Egyptian cat, Felis maniculata. The origin of the striped pattern is easily traced to the European wildcat or to the African wildcat. Of the blotched type Richard Lydekker (Encyclopaedia Brittanica) says: "It may be suggested that the blotched tabby type represents Dr. Nehring’s presumed Chinese element in the cat’s parentage, and that, the missing wild stock may be one of the numerous phases of the leopard-cat (F. bengalensis), in some of which an incipient spiral arrangement of the markings may be noticed on the shoulder.”
The attempt is made by many authors to trace all variations in morphology and color to some wild ancestor. To do so appears to me unnecessary, as all such variations might occur under domestication. The strictly domestic color variations in the cat may be considered maltese, white, white-spotting, yellow, and Siamese dilution. Such variations, if they occur in nature, appear to be blotted out, as they are certainly not characteristic of any wild species. They occur in numerous domestic animals and may be said to be variations by which domestic species ‘mimic’ each other.
[Footnote 4. By the use of the term ‘mimic’ I wish merely to denote what in my estimation underlies many at least of the phenomena which biologists have attempted to explain by the mimicry hypothesis. There are only a limited number of ways in which an organism may vary. Thus a mammalian coat may vary in dilution and distribution of the pigments black, brown, and yellow. No other pigments can be developed. Numerous cases of resemblance, moreover, are in all probability due to homologous factorial differences, even in widely separated species. Metz (’16) has shown that mutations have occurred in Drosophila virilis (species B) producing characters similar to mutant characters in D. ampelophila. Such variations are inherited according to a similar mechanism and show comparable linkage relationships. I am of the opinion that resemblances in the colors and patterns of different mammals are often due to such genetic homology.]
On the other hand, variations in the ticking factors and in the banding or pattern factors occur both in wild and in domestic mammals. Such variations in the domestic cat produce color patterns closely similar to numerous wild species. Variations from red to silver occur in wild cats. The tiger has a high degree of red with a moderate amount of ticking. Thus the pattern is very well marked. In the lion and the puma as well as in the jungle cat and others, the red is reduced to yellow while the ticking is very intense. Hence the pattern appears only in young animals and is obliterated by the increase of ticking incident with maturity. Other cats like the ounce or snow-leopard and Pallas’ cat represent an extreme reduction of yellow pigment comparable with silvering in domestic tabbies. Loss of agouti producing black varieties of leopards and others are well known. Small species of African and Asiatic cats vary so in color that, much confusion has resulted in taxonomy. All of this diversity may apparently be reduced to variations in ticking, in banding, •and in the red-silver series. Spots, I believe, are produced by crossing of longitudinal and transverse waves of pigment-forming metabolic activity. In these respects the domestic cat tends to 'mimic’ its wild relatives, but whether the variations have originated by crossing or by mutation is an open question.
IV. THE SUMMARY
The inheritance of color variations in the domestic cat has been investigated at the Zoological Laboratory of the University of Pennsylvania.
Literature Cited By Whiting (some with relevant quotes included)
Bateson, W. 1913 Mendel's Principles of Heredity. Cambridge University Press.
Castle, W. E. 1916 Genetics and Eugenics. Harvard University Press.
Cutler, D. W., and Doncaster, L. 1915 On the sterility of the tortoiseshell torn cat. Journal of Genetics, December.
Davenport, C. B. 1905 Details in regard to cats. Report on the work of the Station for Experimental Evolution, Cold Spring Harbor.
Doncaster, L. 1905 On the inheritance of tortoiseshell and related colours in cats. Proceedings of the Cambridge Philosophical Society, 13, pt. 1, p. 35.
Doncaster, L. 1912 Sex-limited inheritance in cats. Science, N. S., vol. 36, no. 918, August 2.
Doncaster, L. 1913 On sex-limited inheritance in cats, and its bearing on the sex limited transmission of certain human abnormalities. Journal of Genetics, June.
Doncaster, L. 1914 Chromosomes, heredity, and sex. Quarterly Journal of Microscopical Science. February.
Ibsen, Heman L. 1916 Tricolour inheritance. III. Tortoiseshell cats. Genetics, vol. 1.
Little, C. C. 1912 Preliminary note on the occurrence of a sex-limited character in cats. Science, N. S., vol. 35, no. 907, May 17.
Nehring, a. 1887 Ueber die Sohlenfiirbung am Hinterfusse von Felis catus, F. caligata, F. maniculata und F. domestica. Sitzungs-Berichte der Gesellschaft naturforschender Freunde zu Berlin (On the markings of the soles and hind feet of Felis catus, F. caligata, F. and F. maniculata domestica. Session Reports of Berlin Society of Friends of Natural Science).
PococK, R. I. 1907 On English domestic cats. Proceedings of the Zoological Society of London, Februaiy 5.
Przibram, H. 1908 Vererbungsversuche uber asymmetrische Augenfarbung bei Angorakatzen. Archiv fur Entwicklungsmechanikder Organismen, XXV, S. 260.
Rope, G. T. 1881 On the colour and disposition of markings in the domestic cat. Zoologist, vol. 5, no. 57.
Whiting, P. W. 1915 The tortoiseshell cat. The American Naturalist, vol. 49, August.
Whiting, P. W., 1919 Inheritance of white spotting and other colour characters in cats. Am. Nat., 53 (published after the paper I have summarised).
Williams, Mrs. Leslie. 1908 The cat. Henry Altemus Company, Philadelphia
Wright, Sewall. 1917 Colour inheritance in mammals. The Journal of Heredity, vol. 8.
Chase, Herman B, 1939 Studies On The Tricolour Pattern Of The Guinea Pig. The Distribution Of Black And Yellow As Affected By White Spotting And By Imperfect Dominance In The Tortoise Shell Series Of Alleles
THE TORTOISESHELL TOMCAT — A SUGGESTION. By L. DONCASTER, F.R.S.
Professor of Zoology, Liverpool University.
JOURNAL OF GENETICS, Volume IX. 1919-1920
In the Journal of Genetics, viii. 1919, p. 279, C. C. Little puts forward a new hypothesis to account for the origin of the tortoiseshell tomcat. He suggests that the sterile tortoiseshell male is comparable with the X0 males produced in Drosophila by the fertilization of a non-disjunctional ovum bearing no sex-chromosome by an X-bearing spermatozoon. Such males in Drosophila are sterile, and not infrequently show mosaic characters. Little therefore suggests that the sterile tortoiseshell male is of similar origin, is sterile from the same cause, and is tortoiseshell instead of yellow owing to a tendency to mosaic distribution of the yellow factor. Fertile tortoiseshell males are explained on the assumption that by secondary non -disjunction the X and Y (X and 0) chromosomes come into the zygote from the same parent, and it is suggested that under these circumstances the "0" chromosome does not have its normal effect, and that what would otherwise have been a yellow male becomes a tortoiseshell. For this second assumption there is little or no real evidence, and its author himself seems somewhat diffident about it. The comparison of the sterile tortoiseshell male with the sterile X0 type of male in Drosophila is at first sight more attractive. The comparison, however, really rests only on the fact of sterility, for the tendency to a mosaic distribution of factors does not seem to be comparable in the two cases. In Drosophila, as Morgan, Bridges and Sturtevant show in their recent monograph on Drosophila (7), the mosaic distribution of factors is almost certainly due to abnormal distribution of the X-chromosome in the segmentation of the egg, and the flies are almost always mosaics of sex-characters (gynandromorphs) no less than of inherited factors. The inherited factors accompany the abnormal distribution of the sex-factors, so that different parts of the fly show not only different inherited characters but also different sexual features. There seems to be no reason for supposing that this is so, or for expecting that it should be so, in a tortoiseshell cat of chromosome constitution X0 instead of XY (X- instead of X0).
The purpose of this note is to put forward another suggestion — equally speculative, but more easily capable of verification. In a recent paper Magnusson (6) has described the anatomy, both gross and microscopic, of some 70 freemartins. He finds that although in all of them the external features are predominantly female, internally they approximate more or less nearly to the male. In more than half of them the gonads were in the position of ovaries, the uterus was distinctly developed and the vasa deferentia rudimentary, but even in these the gonad was at least as much like a rudimentary testis in structure as like an ovary. The examples described by Miss Chapin (1) from Lillie's material seem to correspond with this group of Magnusson's cases. In the remainder of his cases Magnusson found a series of stages in which the uterus was more and more reduced down to complete absence, the vasa deferentia and epididymis well developed, and in several instances the gonads had passed into the inguinal canal. In these examples of more male type the testis contained seminal tubules with interstitial tissue, including Leydig's interstitial cells, between them. The tubules sometimes contained Sertoli cells, but never any trace of true seminal cells, and Magnusson describes them as closely similar to those found in retained (cryptorchid) testes of true males. Further, it should be noted that in a number of his examples the interstitial tissue was much more abundant than in normal testes.
Magnusson's description of the microscopic structure of the gonad in the more masculine of his freemartins in Cattle immediately recalls the condition described by D. W. Cutler and the writer (2) in the testis of a sterile tortoiseshell tomcat; the structure of the cat's testis appears to be almost identical with that of the gonad in some of the free-martins. This suggests the possibility that the tortoiseshell tomcat may possibly be in fact a freemartin. Lillie (4) has shown almost beyond doubt that the freemartin is derived from a female embryo which has been "masculized " by the confluence of its vascular system with that of a neighbouring male foetus. Magnusson, not knowing Lillie's work, believes that the freemartin is one of a pair of uniovular male twins, but his evidence for this is entirely unconvincing, and it may be assumed with confidence that Lillie's explanation is the true one. If in Cats, as in Cattle, the embryonic membranes of two foetuses may coalesce so that their blood-system becomes confluent, and if in them the same masculization of a female embryo results, occasionally a tortoiseshell female embryo would be masculized. It must be assumed that the masculization is more complete in the Cat than in Cattle, so that the testes, instead of only comparatively rarely descending into the inguinal canal, regularly reach the scrotum, and the external genitalia also assume the male type. Since, however, Magnusson finds a series of stages between an almost female and a nearly male condition, it is not a very improbable assumption that in the cat the process is habitually carried further. Possibly the fertile tortoiseshell male, which on the evidence of Sir Claud Alexander (quoted by Cutler and Doncaster) must be admitted to exist, is the final stage of the series. Complete conversion into males of embryos which are genetically female appears to exist in some of Goldschmidt's Lymantria crosses (3) and in several hybrids between different species of moths.
This hypothesis involves the assumption that females of colours other than tortoiseshell must also at times be changed into apparent males, but since these would naturally be regarded as males which for some reason or other were not successful breeders, no record of them would be preserved. The suggestion here put forward does not seem difficult to test. By collecting gravid uteri of cats and examining the foetal membranes it should not be difficult to find out whether confluence between the blood-systems of neighbouring foetuses ever takes place. If it did, and if one of the foetuses was definitely male, then according to the hypothesis the other foetus should, in two cases out of three, have testis-like gonads with no seminal cells in the tubules.
The writer does not feel confident of being able to carry out the considerable labour involved in testing the hypothesis in the immediate future, and he puts it forward in the hope that some other may be able to obtain and examine the necessary material.
PAPERS REFERRED TO.
INHERITANCE OF WHITE-SPOTTING AND OTHER COLOR CHARACTERS IN CATS - Dr. P. W. Whiting
The American Naturalist , Vol. LIII. November-December, 1919 No. 629
In a previous paper [Whiting, P. W., "Inheritance of Coat-Color in Cats,” The Journal of Experimental Zoology, Vol. 25, No. 2, April, 1918] I have presented data bearing on the general subject of the inheritance of coat-color in cats. The experiments at the University of Pennsylvania were still in progress when the paper was published and further results have since been obtained. Unfortunately an eczema infected the stock and the investigations were brought to an end by the death of several animals. It is thought advisable therefore to present the rest of the data in the present paper and to summarize results thus far obtained.
Numbers denoting individuals or matings are inserted as in the previous paper for the purpose of cross reference. In the genetic formulae A' denotes much-ticked; A, little-ticked; a, non-ticked; B' denotes lined; B, striped; b, blotched; M, denotes intensely pigmented; m, maltese dilution; W denotes dominant solid white; w, color; Y de-notes yellow; y, black; Yy, tortoiseshell. Symbols are omitted when character was for any reason undetermined.
A cream male (24) (b.m.w.Y) was crossed (48) to a solid-white yellow-eyed half sister (29) (W) from mating 43 (a white male by an "anomalous” (Yy) cream female (23) mentioned below). There were produced one orange male (b.M.w.Y), and one cream female (b.m.w.Y). Both had an extreme amount of white-spotting. The same male (24), when crossed (51) to his "anomalous” (Ty) cream mother, (23) (b.m.w.Yy), sired two cream females (b.m.w.Y). He was also crossed (53) to a black female, (32) (a.B.M), and sired one maltese male, (a.B.m.w.y) and four tortoiseshell females, (one a.B.M, one b.m, and two a.M).
The progeny from the "anomalous” (Yy) cream female (23) by her cream son (24) now consists of four litters (41, 46, 50, and 51) containing two maltese males (yX —), one cream male (YX —) and four cream females (YX IX).
There are now in all eight matings of "yellow” male (YX —) by "black” female (yX yX) giving sixteen "black” males (yX —) and seventeen "tortoiseshell” females (YXyX).
Dr. Charles Penrose, of Philadelphia, very kindly loaned his Caffer cat for crossing, a much-ticked lined male (21) (A'.B'.M.y) mentioned in the previous paper. A cross (52) made with an orange striped female (31) (B.M.Y), from mating 37 produced three tortoiseshell females,—-a much-ticked lined (A'.B'.M.Yy), a lined with ticking present but with so much yellow that the exact degree was uncertain, (B'.M.Yy), a ticked of uncertain degree in which the banding was also uncertain on account of admixture of black and yellow, (M.Yy).
The same male (21), crossed (55) to an orange striped sister (33) (B.M.Y) of female 31, sired five orange lined males (B'.M.Y).
The same male (21) crossed (54) to a tortoiseshell (28) (a.B.M.Yy) sired four lined non-yellow kittens,—two little-ticked males (A.B'.M.y), and two much-ticked females (A'.B'.M.y).
The same male (21) crossed (56) to a blotched maltese tortoiseshell (13) (A.b.m.Yy) sired four lined orange males (B'.M.Y) and two lined tortoiseshell females (B'.M.Yy).
When he was crossed (57) to a dilute tortoiseshell (34) (a.b.m.yY), there were produced a lined orange male (B'M.T) and a lined tortoiseshell female (B'.M.Yy).
When he was crossed (58) to a tortoiseshell female (30) (a.B.M.Yy) he sired two orange lined males (B'.M.Y), one little-ticked, lined male (A.B'.M.y), and two lined females,—one tortoiseshell with so much yellow that degree of ticking could not be made out (B'.M.Yy), and one non-yellow with so much white that degree of ticking could not be made out (B'.M.y).
The crosses of this Caffer cat are reciprocal to those summarized above, crosses of "yellow” males (YX —) by "black” (yX yX) and by "tortoiseshell” (YX yX) females. Here we have a "black” male (yX —) by "yellow” females (YX YX) giving five "yellow” males (YX —) and three "tortoiseshell” females (YX yX) and a "black” male (yX —) by "tortoiseshell” females (YX yX) giving three "black” males (yX —) and seven "yellow” males (YX —) and four "tortoiseshell” females (YX yX) and three "black” females (yX yX). It may be seen, therefore, that the principle of sex-linkage applies in all these cases.
The progeny of the Caffer cat (21) are of interest also from the point of view of ticking and banding. The essential characteristic of Caffer is the narrow banded or "lined” condition. Banding of intermediate width, "striping,” acts as a recessive as previously shown and the widest bands, "blotches,” are recessive to both "lines” and "stripes.” The parents (18, 19) of this cat were both lined, but produced blotched offspring as well as lined. Evidently this cat (21) is the homozygous segregate, for twenty-four of his twenty-five offspring are certainly lined and in the other (52.3) there is so much white-spotting and so much intermixture of yellow and black in the pigmented areas that the condition of banding is uncertain. It is probable, however, that even in this case a wider type of bands,—stripes or blotches, would have been more easily seen. Of the females to which cat 21 was crossed, two (13, 34) were blotched, and the other four (28, 30, 31, 33) were striped but known to be carrying blotched. Lined is therefore dominant to both striped and blotched as previously stated. The results thus far obtained do not demonstrate the allelomorphism of the three types of banding. In order to do that it would be necessary to cross one of the offspring carrying striped (B'B) to blotched cats (bb). All kittens should be lined (B'b) or striped (Bb). If blotched occurred it would demonstrate that two loci were involved, LI and Ss. Blotched would then be ll.ss, and the nomenclature would have to be changed.
The production of orange and tortoiseshell lined cats is of interest. They are as expected in every way comparable to other oranges and tortoiseshells except for the narrower bands.
Results in regard to ticking may now be considered. In the previous paper ‘a’ was used to denote lack of ticking; A, little-ticked or dark tabby; and A', much-ticked or light tabby. It now appears that there are two hereditary grades of ticking previously grouped under A' between which there is a fairly wide difference. Comparison of kittens at birth or of adult cats makes the distinction clear. During growth intergradations appear for ticking increases with age as in rodents. A' should therefore be divided into Ac, extreme-ticking, and Am much-ticking. Fully as much difference exists between Ac and Am as between Am and A.
A blotched male (11) crossed (14) with a black female (15) sired four blotched kittens, and a blotched female (14) crossed (31) with a lined little-ticked male (18) produced one lined and three blotched offspring. The two blotched cats (11 and 14) were extremely-ticked, Ac, as were also the eight kittens. A much-ticked, Am, Caffer female (19) when crossed (19) to the little-ticked, A, Caffer male (18) produced one much-ticked (21) and three little-ticked. The much-ticked mother (19) and son (21) are very similar and contrast strongly with the extremely-ticked cats mentioned as well as with little-ticked. With the exception of five kittens, the offspring of the much-ticked male (21) are useless for determining degree of ticking on account of the presence of yellow. Three kittens (58.2, 54.3, and 54.4) are little-ticked like their grandfather (18). Two kittens (54.1, 54.2) were much-ticked like their father and grandmother. The mothers of all of these kittens were non-ticked. The same degrees of ticking, Am and A, have been possessed by three generations.
Skins illustrating the three types of ticking Ac, Am, and A are preserved for reference.
The crosses summarized in the preceding paper and above may now be considered from the point of view of white-spotting. Solid-white acts as a complete dominant to other colors as shown in the previous paper. White- spotting as seen among cats in general grades all the way from solid-white to self. In individual fraternities, however, it may show wide and clean segregation as the crosses below demonstrate. A "self” cat may have a minute white spot on breast of belly or a few sparsely scattered white hairs. In this case it might be called near-self. Restricted spotting denotes white on nose, breast, belly, or feet. It segregates widely from near-self in the crosses here considered, but grades into moderate spotting, which denotes the further extension of white to sides of body as well. Moderate spotting in turn grades into considerable, which denotes more white than color. Extreme spotting denotes that pigment is limited to small spots on head, back, or tail.
Crosses involving only self, restricted and moderate spotting, and solid-white may be considered first.
A self male (18) crossed (19, 31) to two self females (19, 14) sired eight self. One of these self offspring (21) crossed (55) to a self female (33) sired five self. Self may therefore breed true.
The first mentioned self male- (18) crossed (28, 29) with two restricted spot females (10, 2) sired three self and four restricted spot. The other self male (21) crossed (54, 56) to two restricted spot females (28, 13) sired five self and five restricted spot. A restricted spot male (8) crossed twice (9, 30) to a self female (20) sired three self and seven restricted spot. A restricted spot female (3) crossed (16) to a self male (6) produced six self. Self by restricted spot therefore has produced seventeen self and sixteen restricted spot, the expectation if restricted spot is heterozygous.
The restricted spot male (8) crossed (12, 33) to two restricted spot females (25, 3) sired one self, three re-stricted, and one moderate. This is in line with expectation if spotting is dominant, the moderate in this case possibly representing the homozygote.
The same restricted spot male (8) crossed (32) to a solid-white (22) sired two solid-white and two completely self. This is in line with the assumption that the white female was homozygous for self, ss, and heterozygous for color, Ww, or that spotting and white are both allelomorphic with self and that she was carrying self. The male would then be w1w, w1 standing for restricted or moderate spotting.
Crosses involving greater amounts of spotting may now be considered.
The self male (21) crossed (52) to a considerable spot female (31) sired one self and two considerable. When crossed (58) to a moderate spot (30) he sired three self and two considerable. These results show that considerable segregates from self and that a greater degree of spotting may be produced from a less by crossing to self. Modifiers are indicated.
The same male was also crossed (57) to a considerable (34) and sired two restricted. In this case modifiers may have been assorted to produce restriction, but the female (34) was derived from a cross (34) of a considerable (24) by a restricted (3) each of which was known to carry self. She may therefore have been of composition wm, much spotting, being derived from her considerable parent and w1, little spotting, from her restricted parent. She could then produce restricted offspring, w1w, when crossed to self.
Offspring of self by spotted known to carry self are therefore twenty-one self and twenty spotted.
The considerable spot male (24) was crossed to various spotted females known by these or other crosses to produce self. With a restricted (3) he sired (34, 49) two self, one restricted, three moderate, and three considerable. With a moderate (32) he sired (53) two self, one restricted, one moderate and one extreme. With a restricted (28) he sired (44) one self, one restricted, one moderate, and one considerable. With a moderate (30) he sired (45) two considerable and one extreme. With a restricted (13) he sired (47) two self, two considerable (one of which graded toward extreme), and one very extreme. This last cross is interesting for the offspring vary far in both directions from the parental types.
Crosses of spotted by spotted when both carry self have produced twenty-three spotted to eight self which is very close to the three-to-one expectation.
The considerable spot male (24) above mentioned when crossed (41, 46, 50, 51) to his considerable spot mother (23) sired five considerable and two restricted, the segre-gation being striking through failure of any moderates to appear. This is in line with the supposition that the mother (23) was carrying little spotting and was therefore of composition tv'"tv1. The cross might therefore be wmw x wmw1 = 5(wmwm, wmw or wmw1) + 2(w1w).
The same male (24) (wmw) was crossed (48) to a solid- white half sister (29) from the same mother (23) (wmw1) by a white male (W ?). There were produced two extreme spot. The white female (29) may therefore have been Wwm and the extreme spot offspring wmwm.
The failure of anything higher than restricted spotting to occur among the offspring of restricted by self, although cats with considerable may carry self, indicates that there may be allelomorphic factors determining different degrees of spotting. In any case it appears that self is recessive to spotting and that color is recessive to solid-white. The principle is suggested that there is a quadruple allelomorphic series:— W, solid- white; wm, much spotted; w1, little spotted; and w, self, with dominance in the degree of decreasing pigmentation. Crosses of white to self and of spotted to self would be of value in checking this principle. Any one white cat might throw besides white either much spotted, little spotted, or self; a much spotted might throw besides much, either little or self, and little should throw little or little and self. If three distinct types were produced from any one white or spotted cat crossed to numerous self cats, this would demonstrate modifiers of considerable importance or disprove the hypothesis of allelomorphism suggested.
Attention should be called to an interesting but unex-plained relation that exists between yellow- and white- spotting. "Self” tortoiseshells have yellow hairs closely intermixed with non-yellow. This makes it very difficult to determine degree of ticking in such animals. Tortoiseshells with restricted white-spotting tend to have yellow separated into patches, while further extension of white separates yellow and non-yellow areas still more. Separation of yellow into patches appears not to be correlated with amount of yellow.
GENERAL SUMMARY OF INHERITANCE OF COAT-COLOR IN CATS
It may be of interest to summarize very briefly the genetic data thus far collected on coat-color in cats. Ratios are not significant since fraternities from homo-zygous dominants and heterozvgotes are included together.
Maltese dilution, m, is presumably a simple recessive to intensity, M. Intense by intense have produced 41 intense. Intense by dilute have produced 37 intense and 23 dilute. Dilute by dilute have produced 18 dilute.
Solid-white, W, evidently acts as a simple dominant over color, w. It is true-breeding in the hands of fanciers. White by color (amount of white-spotting undetermined) have produced 3 white and 4 colored (one near-self). Table I shows summaries for white and white-spotting of determined degree. It is obvious that although extensively pigmented animals appear among the offspring of cats showing much white there is little tendency for a kitten to show more white than appears in either parent. Table II gives a summary of the results thus far collected in reference to the inheritance of yellow. Doncaster’s [Doncaster, L., "On Sex-limited Inheritance in Cats, and Its Bearing on the Sex-limited Transmission of Certain Human Abnormalities,” Journal of Genetics, June, 1913.] summaries from fancy breeders and from Little’s data are given, kittens of undetermined sex being omitted. The three tortoiseshell females from one pair of Doncaster’s yellow by yellow may be readily explained if it be supposed that the mother was an extreme yellow variant of the heterozygote Yy, comparable with my cream female number 23. Anomalous black females may be similarly heterozygous. Anomalous blacks and tor-toiseshells are to be expected from anomalous yellow females. Anomalous offspring are recorded in italics in Table II.
As regards banding, certain creams and blacks could not be classified and are consequently omitted from the summaries. Lined by lined have given 2 lined and 2 blotched. Lined by striped have given 17 lined and 2 striped. Lined by blotched have given 12 lined and 4 blotched. Striped by blotched have given 19 striped and 8 blotched. Blotched by blotched have given 4 blotched.
As regards ticking, it is necessary to omit all yellows and many tortoiseshells, as well as some with much white. Extremely-ticked by little-ticked have given 4 extremely- ticked. Extremely-ticked by black have given 4 extremely-ticked. Much-ticked by little-ticked have given 1 much-ticked and 3 little-ticked. Much-ticked by black have given 2 much-ticked and 3 little-ticked. Little-ticked by little-ticked have given 5 little-ticked and 1 black. Little-ticked by black have given 7 little-ticked.
COLOUR INHERITANCE IN CATS, WITH SPECIAL REFERENCE TO THE COLOURS BLACK, YELLOW AND TORTOISE-SHELL. BY C. C. Little 1919.
JOURNAL OF GENETICS, Volume VIII. 1918—1919
Research Associate. Carnegie Station for Experimental Evolution
(Cold Spring Harbor, New York).
I. INTRODUCTORY.
This paper has two objects: (1) the critical examination of experimental data on, and of current hypotheses concerning the inheritance of black, yellow, and tortoise-shell coat colours in cats; (2) the suggestion of possible explanations for the occurrence of (a) unexpected colour classes in ordinary crosses between blacks, yellows, and tortoise-shells, and of (b) both sterile and fertile tortoise-shell males which appear extremely rarely.
The fact that the work of all investigators of this subject has left the two points above mentioned not satisfactorily accounted for justifies an attempt to explain the observed experimental results, even though at this time no additional breeding data are offered for consideration.
II. THE FACTS REQUIRING EXPLANATION.
The critical and apparently contradictory facts which have been brought out by breeding experiments with cats, and which must be satisfactorily accounted for and explained, are briefly as follows:
(1) In crosses between yellow males and black females, where the expectation on the basis of complete sex-linkage is black males and tortoise-shell females, black females are sometimes produced. (Doncaster, 1913.)
(2) In crosses between yellow males and tortoise-shell females, where yellow males, black males, yellow females and tortoise-shell females are the only classes expected on the basis of black females are sometimes produced. (Doncaster, 1913.)
(3) In crosses between two yellow cats, although only yellow young are expected, two aberrant results have been noted.
(3a) A mating of this type has produced tortoise-shell females besides yellows of both sexes. (Doncaster, 1913.)
(3b) A mating of this type has produced tortoise-shell females and black males besides yellows of both sexes. (Whiting, 1918.)
(4) There is no record of two black cats crossed together having given yellow or tortoise-shell young.
(5) Tortoise-shell males are produced much more rarely than any of the aberrant classes recorded under headings 1, 2, and 3 above. (Doncaster, 1913; Wright, 1918.)
(6) Such tortoise-shell males are usually sterile. (Cutler and Doncaster, 1915.)
(7) If they are not sterile they apparently do not give tortoise-shell sons, but breed as yellows. (Doncaster, 1913.)
In considering these facts, investigators have usually tried to explain all of them by a single hypothesis. (Doncaster, 1913; Whiting, 1918.) This has proved to be difficult and unsatisfactory. (Ibsen, 1916 ; Wright 1918.) It is believed that the experimental evidence favours the existence of two genetically independent agents at work in the production of these aberrances, for
(a) The appearance of the unexpected individuals noted under headings 1, 2, and 3 above, is relatively frequent, and produces regular results involving neither sterility nor the formation of new colour types.
{b) On the other hand, the occurrence of tortoise-shell males is very infrequent, not regular, and is in a majority of cases intimately connected with sterility.
Such being the case, an effort will be made to explain the appearance of the unexpected individuals noted under headings 1, 2, and 3 by one hypothesis and the occurrence of tortoise-shell males by a different one.
III. THE RELATION BETWEEN YELLOW AND BLACK.
One of the first points to be established is the nature of the genetic relation between yellow coat colour and black coat colour. In this connection Ibsen, 1916, and Wright, 1918, believe black or extension of black pigment to the coat, to be epistatic to yellow or the restriction of black pigment from the coat. Doncaster, 1913, and Whiting, 1918, consider the two coat colours allelomorphic, the heterozygote being commonly tortoise-shell. The terminology used by them is as follows:
Ibsen, 1916: Black B is dominant to orange b which is borne in the X chromosome. Under ordinary conditions the factor for orange b is closely linked to T, a factor for tortoise-shell which acts only in the presence of B-black. The female is XX, the male is X0 in formula.
Wright, 1918: Black is due to the action of a factor A, while tortoise-shell is produced by heterozygosis of an "extension” factor E. Tortoise-shell females are thus Ee, yellow males e-, and black males E-, in formula. The factor E is borne in the X chromosome. The female is XX, the male X0 in formula.
Doncaster, 1913 considers that yellow and black are allelomorphic, and expresses yellow by Y, and black by B. Where both are present, a YB or tortoise-shell animal is produced. The female is XX, the male X0 in formula.
Whiting, 1918 also considers yellow Y to be allelomorphic with black y, and supposes y to be borne in the X chromosome. The female is homozygous, the male heterozygous for X.
In 1912 I employed much the same terminology as that of Doncaster, but in view of the production of blacks and tortoise-shells by two yellows and the failure of blacks when crossed inter se to produce anything except blacks, it is probable that the relationship between these two colours may be more accurately expressed in somewhat the following manner: B a factor for the production of black pigment which is found in all X gametes. Y a factor for the restriction of black pigment from the coat allelomorphic to y, a factor for the extension of black pigment to the coat. One " dose " of Y is normally completely epistatic to one "dose" of B, thus producing yellow individuals; but two "doses" of B to one of Y produces a tortoise-shell. The factor Y and its allelomorph y are also borne in the X chromosome. Thus:
YBX YBX Yellow female
YBX 0 Yellow male
yBX yBX Black female
yBX 0 Black male
YBX yBX Tortoise-shell female
This type of relationship will become clear us the crosses are taken up in detail, and is further made use of in explaining the occurrence of tortoise-shell males.
IV. AN ATTEMPT TO EXPLAIN THE APPEARANCE OF UNEXPECTED INDIVIDUALS OF NORMAL COLOUR TYPES. (HEADINGS 1, 2, AND 3, SECTION II, ABOVE.)
It is tacitly assumed by all investigators that at some time or times in the past, there must have been a genetic change, ridding certain gametes of the epistatic colour factor, whether it be the Y of Whiting, the E of Wright, or the T of Ibsen. Had this not been the case neither the hypostatic form nor the tortoise-shell heterozygote could have appeared. We may, then, for the sake of argument accept the set of symbols given above, and assume that the change from Y to y must have occurred. There is no experimental evidence to show how recently or how frequently this change may have taken place, but if we assume that it is still taking place in a portion of the gametes of certain individuals — which seems entirely probable — all the results obtained under headings 1, 2, and 3, may be accounted for. Such a change from an epistatic to a hypostatic condition would be directly comparable to the appearance of the recessive pink-eyed mutation in a stock of dilute brown mice recorded by the writer in 1916. Animals in whose gametes this mutative process was occurring de novo would show no trace of it in their own somatic characteristics, but would, upon breeding, give results in agreement with the actual aberrant classes obtained.
We should thus expect that an occasional yellow female would form gametes yBX in addition to those containing YBX which she normally produces. Similarly, certain yellow males would be found which showed by their progeny that they were forming among their X gametes some which were of the constitution yBX instead of the normal YBX type. Yellow males of this unusual kind would, when crossed with black females, give among their progeny a certain number of black females, in number depending upon the frequency with which the unusual yBX sperm was formed. This fact would explain the aberrances listed above under Section II. Heading 1.
Similarly, such unusual yellow males would, when mated to normal tortoise-shell females, give rise to a certain number of black females in addition to the other classes normally expected. This would cover category two of exceptions mentioned above (Section II). Finally, a yellow forming yBX gametes, when crossed with a normal yellow or with one of its own type, would give rise to unexpected black or tortoise-shell young, the proportion depending upon whether the yellow male of the female or both were concerned in the formation of the yBX gametes.
Thus if the male was alone concerned, tortoise-shell females, but no black males would be likely to appear among the progeny. This appears to be the case in the mating recorded by Doncaster (1913) in which two yellows gave among their progeny three blue females with a cream coloured patch (tortoise-shells). If, on the other hand, the female parent was the unusual mutative individual, black males would occur in addition to tortoise-shell females and yellows of both sexes. This condition was realised in the case of female dilute yellow #23 (formerly owned by me) whose breeding record is reported by Whiting, 1918. An explanation of this sort would account for the aberrances noted under Section II, Heading 3, above.
From the number of tortoise-shell and black young obtained in the two cases referred to, and from the numerical relation of the black females under headings 1 and 2 (Section II) to the expected colour classes (Doncaster, 1913), it seems probable that yellow animals forming yBX gametes do so in approximately 50% of the gametes they form, as would a normal heterozygote.
In addition to yellow animals, certain tortoise-shell females might theoretically be expected to show the same phenomenon. Such animals would form an excess of, or possibly exclusively, yBX gametes, and, in so far as they did so, would breed as blacks. Such an occurrence would, however, give rise to no unexpected classes of young in crosses, but might result in the absence of some of those normally expected from certain matings. Quite naturally this fact might, in a small number of progeny, escape notice.
There is no evidence to show that the appearance of any of the classes above referred to is in any way connected with a break in sex-linkage or with the occurrence of tortoise-shell males, and we may therefore, until such evidence is presented, fairly consider them as independently produced.
V. CRITICISM OF EXISTING HYPOTHESES TO EXPLAIN THE APPEARANCE OF UNEXPECTED INDIVIDUALS OF NORMAL COLOUR TYPES. (HEADINGS 1, 2, AND 3, SECTION II, ABOVE.)
Attempts to explain the appearance of the aberrant colour classes referred to, have involved either (a) the breaking of sex-linkage with " crossing over " in the male, or (b) the occurrence of a series of modifying factors determining the relative degree of black and yellow pigmentation. They may be separately considered as follows :
(a) Doncaster's hypothesis of a break in sex-linkage : this hypothesis, which in a modified form is a basis for Ibsen's later explanation of the appearance of unusual colour types, involves, if it is to explain the exceptional black females, the existence of "crossing over" in the male between the X and the 0 chromosomes. Such crossing over has not, in so far as I am aware, been demonstrated in any forms in which the male is X0 in formula as in cats. It further would suppose that, as tortoise-shell males were formed by the same process, they would be expected to occur with as great frequency as the exceptional black females. It further leaves entirely unexplained the appearance of blacks or tortoise-shells from a cross between yellow animals. These objections seem to be of sufficient weight to throw the chances against Doncaster's or Ibsen's hypotheses.
(6) Whiting's hypothesis of modifying factors which at one end of the series would serve to make tortoise-shell animals yellow, and at the other end of the series make them black, remains as a possibility though seriously invalidated by certain points as follows :
(b1) There should be records of black females (genotypically tortoise-shell) which if crossed with other blacks should give yellow males and tortoise-shell females, or if crossed with yellow males should give unexpected yellow females. Neither of these results has been recorded.
(b2) Doncaster, 1913, reports that the three tortoise-shell females produced from a single cross between two yellows were " blue with a cream patch " thus showing that they were near the black end of Whiting's modifier series. Inasmuch as under his hypothesis one of their parents must have been at the opposite or yellow end of the series, it is difficult to explain how and why many of its progeny should show the condition characterising nearly the other end of a graded series.
(b3) The occurrence of these young in a single mating makes it seem likely that the particular animal was forming ordinary yBX gametes in a considerable number.
(b4) The tortoise-shell young produced by dilute yellow female #23 already referred to, before she was sent to Dr Whiting, were normal tortoise-shell in colour ; if anything, more nearly on the black end of the graded series, than on the yellow. This case serves to support that reported by Doncaster, and tends to show that the yellow animal transmitted the its progeny no peculiar set of modifiers.
VI. AN ATTEMPT TO EXPLAIN THE OCCURRENCE OF (a) STERILE, AND (b) FERTILE TORTOISE-SHELL MALES
(a) The production of sterile tortoise-shell males.
It is agreed by all those who have reported on breeding experiments with cats that the female appears to by homozygous, the male heterozygous, for sex. The former is therefore XX, the latter X0 in formula. This places cats in the same category with Drosophila, and this in turn means that one may rightfully turn, and in fact should turn, to the magnificent work of Morgan and his associates in any attempt at explaining a peculiar result which shows exceptional conditions of sex-linkage.
If one considers the phenomena of non-disjunction of the X chromosome in Drosophila, reported by Bridges in 1913, and later (1916 a and b) further established by him after an extensive series of breeding experiments, one cannot fail to be impressed by the similarity between the results of that process in Drosophila, and the observed experimental facts in cats. For example, non-disjunction is neither frequent in its occurrence nor is it clear enough in its hereditary behaviour to give striking numerical results in as slow breeding an animal as a cat, unless it were watched for deliberately. In Drosophila it gives rise to two very significant exceptions to the normal sex-linked inheritance. First, it produces animals apparently males, which are sterile, and second, mosaic forms are apt to arise in non-disjunctional stocks. If one considers that the majority of tortoise-shell cats which are apparently males are sterile, and second that they are also a mosaic form in a sex where commonly none is found, the comparison becomes interesting and extremely suggestive.
We may now consider what the probable results of non-disjunction would be, did this phenomenon exist in cats. The characteristic of primary non-disjunction is that in oogenesis the two X chromosomes go together into a single egg, leaving another egg without even the normal single X. This may be shown as follows :
Non-disjunctional female XX forms gametes XX and "-”
If now the eggs of such a female are fertilized by sperm of a normal male we have four possible types of zygotes.
Eggs XX, Sperm X, Zygote XXX = Dies
Eggs "-”, Sperm X, Zygote X”-” = "Near male" always sterile
Eggs XX, Sperm 0, Zygote XX0 = Female with peculiar gametic condition
Eggs d, Sperm 0, Zygote 0”-” = Dies
Bridges has demonstrated that the XXX and 0”-” forms die, and that the X”-” form although appearing like a male is always sterile. If now we imagine a cross to be made between a tortoise-shell female cat showing non-disjunction and a normal yellow male, we should have the following condition :
Non-disjunctional Tortoise-shell female YBXyBX forming gametes YBXyBX and "-”
Normal Yellow male YBX0 Forming gametes YBX and 0
Zygotes
(a) YBXyBYBX = Dies
(b) YBXyBX0 = Tortoise-shell with peculiar gametic conditions
(c ) YBX”-” = "Near male" always sterile
(d) "-”0 = Dies
If now one assumes that absence of the 06 chromosome allows the "near male" class (c ) to develop into a tortoise-shell, disturbing the normal relation of yellow to black to produce a somatic mosaic, we could account for the appearance at rare intervals of tortoise-shell "near males" which were not fertile. It seems not unlikely that the absence of the 0 chromosome might well upset the somatic relationships of certain of the characters whose factors are carried by the X chromosome. This would account for the appearance of a tortoise-shell "male" from a mating of yellow male x tortoise-shell female. (Doncaster, 1913.)
Another mating which, according to Doncaster, has produced a tortoise-shell male is that of yellow male by black female. Here, if the black female showed non-disjunction, the following condition would be found :
Black non-disjunctional female yBXyBX forming gametes yBXyBX and "-”
Normal yellow male YBX0 forming gametes YBX and 0
Zygotes
(a) yBX yBX YBX = Dies
(b) yBX yBX 0 = Black female with peculiar gametic conditions
(c ) "-” YBX = Tortoise-shell? "near male" always sterile (as in previous mating)
(d) "-" 0 = Dies
The third type of mating reported by Doncaster as having. produced a tortoise-shell male is that of black male with tortoise-shell female. Here everyone is in difficulty. If, as Doncaster suggests, the occasional crossing over of Y, the factor for yellow, to a 0 gamete is responsible for the production of a tortoise-shell male, nothing that could happen in either the gametes of the black male or of the tortoise-shell female would produce a tortoise-shell male. On Whiting’s hypothesis we should have to suppose that the tortoise-shell female, although she herself showed no marked modifiers (or she would have been black) transmits unusually heavy modifiers to her sons. These gametes would in turn have to be met by an equally heavy set of modifiers from the black male, or a yellow would result.
Further than this, by Whiting's hypothesis the yellow male is YX0 in constitution, and this makes the source of the black that he must produce somatically under the influence of modifiers in order to become a tortoise-shell uncertain. This condition is, of course, not impossible but is highly improbably. Finally, the phenomenon of non-disjunction meets with distinct difficulties. Unless the black male forms gametes with neither the X nor 0 chromosomes present it would be hard to see how the tortoise-shell male could be produced by this mating. Formation of sperm without X or 0 would not be likely. Yet the possibility exists and may therefore be considered. What seems to me altogether more likely is that the breeder's records on which Doncaster based his observation were in this case uncertain or incorrect, a circumstance quite possible in cats even with the best possible intentions.
(b) The production of fertile tortoise-shell males.
We have seen that peculiar tortoise-shell females of formula YBXyBX0 may possibly be produced by primary non-disjunction. If now one of these females is crossed with a black or a yellow male peculiar yellow males of the constitution YBX00 would be formed as follows:
Non -disjunctional Tortoise-shell female YBXyBX0 forming gametes YBXyBX, YBX0, YBX, 0, yBX0 and YBX
crossed with Yellow male YBX0 forming gametes YBX and 0
Zygotes
(a) YBX yBX YBX = Dies
(b) YBX 0 YBX = Peculiar yellow female
(c ) YBX YBX = Yellow female
(d) 0 YBX = Yellow male
(e) YBX yBX 0 = Peculiar tortoise-shell female
(f) YBX 00 = Peculiar yellow male **
(g) YBX 0 = Yellow male
(h) 00 = Dies
(i) yBX 0 YBX = Peculiar tortoise-shell female
(j) yBX 00 = Peculiar black male
(k) yBX YBX = Tortoise-shell female
(l) yBX 0 = Black male
If now such a peculiar yellow **, YBX 00, is mated with any female showing primary non-disjunction — an animal which might well prove to be a fertile tortoise-shell male would be produced. Thus:
Non-disjunctional Black female yBXyBX forming gametes yBXyBX and "-”
Crossed with Non-disjunctional Yellow male YBX00 forming gametes YBX0, YBX, 0 and 00
(a) yBXyBX YBX0 = Dies
(b) yBXyBX YBX = Dies
(c ) yBXyBX 0 = Peculiar black female
(d) yBXyBX 00 = Peculiar black female?
(e) "-" YBX0 = "Tortoise-shell male” fertile ? **
(f) "-” YBX = "Tortoise-shell ' near male ' sterile” ?
(g) – 0 = Dies
(h) – 00 = Dies
Here the assumption is made that an animal formed from the combination of gametes, YBX0 and "-”, may he somatically a tortoise-shell, and that the 6 chromosome which is brought into the zygote by an X-bearing gamete does not in all cases exert its full influence until gametogenesis. The YBX0-male would then be supposed to develop somatically just as does the YBX-animal, but upon gametogenesis the 0 chromosome of the YBX0-male is able to prevent the sterility which exists in its absence. This seems quite possible, for it appears that in Drosophila the 0 chromosome is not needed for the development of the normal male somatic characters, but that it is necessary, however, for successful gametogenesis in the male.
A fertile tortoise-shell male would, when he formed gametes, behave exactly like a normal yellow male. That is to say, although he was himself the product of a combination of X0 and "-” gametes, he would in gametogenesis form only X and 0 gametes, just as would a normal male. This has been the breeding behaviour of the one recorded certainly fertile tortoise-shell male (see Doncaster, 1913) which acted in crosses with tortoise-shell females apparently exactly as a yellow male would have done.
It will be seen that the above hypothesis, although somewhat complicated, is nevertheless in accordance with experimental facts and accounts for sterile and fertile types of tortoise-shell males ; it explains their infrequency of appearance, and possibly their failure to transmit their own colour pattern to their descendants ; it is supported by the work of Bridges with Drosophila — the most completely investigated form showing a similar type of sex-linkage ; it is further capable of experimental tests.
VII. CRITICISMS OF EXISTING HYPOTHESES TO EXPLAIN THE OCCURRENCE OF TORTOISE-SHELL MALES
(1) Doncaster’s hypothesis, as already pointed out, requires "crossing over” in the male between X and 0 chromosomes – a condition not shown to exist in gametogenesis of any X0 male form. It further fails to account for (a) the comparative infrequency of tortoise-shell males as compared with aberrant black females, (b) the sterility of the majority of tortoise-shell males, and (c ) their peculiar behaviour in breeding.
(2) Ibsen's hypothesis does away with the need of crossing over in the male, but fails , as does Doncaster’s hypothesis, to meet of explain points (a), (b), or (c ) stated above.
(3) Whiting’s hypothesis of modifiers would not be able to give a tortoise-shell male which according to his formula would be XY0 without adding a factor for black to the formula given by him to be carried in the X gamete. It further would suppose that by selection (which undoubtedly has occurred) it would be possible to transmit the necessary modifiers to a considerable number of his male progeny, thus forming tortoise-shell males — and this, though great efforts have been made, has proved impossible. Whiting's hypothesis, like those of Doncaster and of Ibsen, takes no account of the sterility of the majority of tortoise-shell males.
(4) Wright's hypothesis is that tortoise-shell males are really XX individuals in which the abnormality lies not in the colour but in the sex. He likens them to certain sex intergrades already described in some forms by other investigators. This hypothesis meets trouble when a fertile tortoise-shell male is encountered. It also is contrary to the evidence obtained by Bridges who shows that in Drosophila XX forms are females, even though they contain other abnormalities of chromosome distribution.
VIII. SUMMARY AND CONCLUSIONS.
(1) The genetic constitution of the normal colour varieties of cat as regards yellow and black pigmentation appears to be as follows: B = a factor producing black pigmentation, Y = a factor which restricts black from the coat, y = a factor allelomorphic to Y and hypostatic to it, allowing black pigment to extend to the coat.
YBX YBX - Yellow female
YBX 0 - Yellow male
yBX yBX - Black female
yBX 0 - Black male
YBX yBX - Tortoise-shell female
(2) The unexpected but normally pigmented individuals appearing in certain matings can be accounted for by supposing that Y becomes y in a certain proportion of the gametes of exceptional individuals.
(3) Sterile tortoise-shell males may possibly be" near males" formed as a result of non-disjunction of the X chromosome and therefore YBX in constitution.
(4) Fertile tortoise-shell males may also be the product of nondisjunction (secondary) and would be zygotes formed from the fusion of gametes YBX0 and "-". These males in gametogenesis would behave as ordinary yellows.
CROSSES WITH SIAMESE CATS. By K. Tjebbes 1924.
Journal of Genetics, December 1924, Volume 14, Issue 3, pp 355–366
(With One Plate.)
Introduction.
Many years ago Bateson (1909) drew attention to Siamese Cats, recommendiug these peculiar animals as subjects for genetical investigations. Bateson supposed them to represent an analogue to the well-known Himalayan rabbits. From private communication with Mr Bateson I learned that Miss Durham and Mr Backhouse had gathered some evidence about the genetics of Siamese cats, but I have not heard of any further experiments, and, as far as I know, nothing was published.
After haviug worked on Siamese and other cats for some years I happened to show the short records of my results to Mr Bateson, who encouraged me to publish them, meagre as they are. Their incompleteness is chiefly due to the fact, that Siamese cats are rather exacting animals. They do not like being out of doors, they cannot be kept in pens and will not thrive without the company of man. They need the cosiness and warmth of a human dwelling and must be treated as pet animals. The females are shy and mating is often difficult. Further; the greater part of my cats are the offspring from a cross between white Persian and Siamese. The white Persians are perhaps still more difficult to breed than the Siamese. The weakness of these cats is very striking and their females are utterly bad mothers; they often eat their kittens at birth or starve them to death a few days after, being wearied of their nursing duties.
[Note, 2018: Blue-eyed white females are often deaf and cannot hear their kittens crying to be fed.]
These difficulties and the high costs of the experiment may account for the small number of individuals (56 cats) upon which the following report is based.
Description of Materials.
The two Siamese cats used in my experiments, No. 2 (male) and No. 19 (female), descend both from one pair of animals imported from Bangkok by a Dutch fancier about ten years ago. The strain has always been kept pure and no variability has ever been noticed except slight differences in the intensity of the pigmentation. The dark parts of the fur (face, ears, feet, tail) are from darkest chocolate to almost black; the lighter parts may vary from a pale greyish buff to a smoky fawn colour. The adult males are considerably darker than either the females or the youug animals. At birth the kittens are white or nearly white; after ten or twelve days the pigment begins to develop. The iris of the eyes is blue, but not sky-blue as in blue-eyed white cats; in the Siamese eye there is always a shade of milky grey in the blue of the iris. The deeper parts of the eye are pigmentless.
The white Persian female, No. 1, belongs to a strain that during many years was kept by a fancier in my district. It was a very uniform and beautiful strain as far as the long silky fur is considered, but some individuals showed dissimilarity of the eyes, the left eye having a different iris colour (yellow) from the right eye (blue). The female used in my first cross was not affected in this way, both irides being sky-blue. Nearly all the members of this family showed somatic or mental defects (sterility, deafness, dirtiness, inability to withstand the simplest difficulties of a cat’s life). This caused much trouble in the course of my experiments. It is to be well understood, that the white Persians, even those with two blue eyes, are not true albinos, but coloured cats, in which pigmentation has been suppressed but for a very slight trace. Most of them have a little black spot — often only a few black hairs — on the neck, the occiput, or the front; and also in the eyes, iris as well as fundus, some pigment is left.
The tabby male, No. 16, belongs to the most common type of the ordinary house cat in Holland. The type is in this country called "Cyprian.” It is characterized by longitudinal stripes of a yellowish grey on a black coat. On the legs and the tail the stripes tun transversely. On the ears and the feet no stripes are visible, but the pigment in those parts is paler than on the back and approaches the colour of the stripes. The pattem is obviously the same as that of so many wild Felidae; it is variable in the details, but the dark ground is always preponderant; the stripes are in all cases narrower than the dark parts between them, in a ratio varying from about 40:100 to 60:100. The intensity of the pigmentation is variable too, and two chief types may be distinguished: a dark type (yellowish grey stripes on pure black) and a pale type (pale bluish grey stripes on dark lead grey). The pale behaves as a unifactorial recessive to the dark and is obviously a dilute form, comparable with the "blue” cats in their relation to the "black” ones.
[In coat color, the presence of a chromogen factor A is assumed, which in Siamese cats is replaced by a recessive allelomorph Ui which causes a reduction of the intensity of pigment when a factor B for chocolate black pigment is present. A dominant factor D carried by the Persian whites suppresses all pigment. A factor G, representing a series of allelomorphs, produces the tabby pattern. G was also carried by the white Persians. A new type of striped
Siamese was produced in the F2 of the Siamese x tabby cross. Long hair was recessive to short hair. The eye colors were not so clear in their mode of inheritance. The production of differences in the color of the two eyes of the same individuals, offspring of the white Persian female parent, was an important finding. Both coat and eye pigmentations were assumed to be modified by quantitative physiological processes.]
DOMINANT BLACK IN CATS AND ITS BEARING ON THE QUESTION OF THE TORTOISESHELL MALES. By K. Tjebbes and Chr. Wreidt. 1926.
Existence of two different kinds of black colour in cats had been indicated in Tjebbes' experiments on Siamese cats, published in 1924. The occurrence of this dominant black in cats was of special interest because it might explain some tortoiseshell males. A mating between Siamese and tabby (wild type) apparently gave 3 blacks. Three other Siamese x tabby matings gave F1 black heterozygotes only, totalling 10 uniformly black coloured offspring.
The F2 generation produced from those black cats produced 4 blacks, 1 tabby and 2 Siamese [refers to pattern, not breed]. The Siamese were genetically blacks; Tjebbes had previously shown that [black] tabby may be combined with the recessive factor that causes the Siamese pattern. An F2 Siamese x an F1 black male gave 1 black, 1 Siamese and 1 tabby. Together, those matings gave 8 blacks (uniform blacks plus Siamese which are genetically black) against 2 tabbies. The data was admittedly scanty, but Tjebbes was aware of dominant black in rabbits, dogs and swine so he contended that cats had both dominant and recessive black mutations. Punnett (1912) proved the existence of dominant black in an acromelanistic Himalayan rabbit, considered the nearest parallel to the Siamese cats.
Tjebbes then mated a Siamese female with a striped yellow male resulting in 1 female tortoiseshell and 2 male tortoiseshells. All earlier workers on tortoiseshell cats had agreed on the extreme rarity of tortoiseshell males. Doncaster encountered only 3 such males amongst 225 males from mating black x yellow, tortoiseshell x yellow, and tortoiseshell x black.
It was believed that all tortoiseshell males were infertile, which led to a variety of hypotheses. It was supposed that they were due to either non-disjunction or to a process equivalent to freemartins in cattle. However, some tortoiseshell males had proven fertile. In the light of the Tjebbes 2 tortoiseshell males produced by a mating of [supposedly] dominant black x yellow, he thought that the co-operation of dominant black with yellow could produce tortoiseshell tom-cats.
Regarding chromosomes, it wasn’t possible to give a full explanation, but Tjebbes mentioned that studies by other researchers indicated that genes were found on the Y-chromosomes of vertebrates. Sex-linked inheritance in mammals had been found in only three cases, making it likely that sex-linked inheritance was often masked by the presence of genes on the Y-chromosome and also that crossing-over between X and Y chromosomes might occur.
DOMINANT BLACK IN CATS AND ITS BEARING ON THE QUESTION OF THE TORTOISESHELL MALES--A CRITICISM. By Ruth C. Bamber (Mrs Bisbee) and E. Catherine Herdman. 1927
Journal of genetics xviii, Vol 18, No. 2, p. 219, 1924.
In the Journal of Genetics for October 1926, vol. xvii, no. 2, pp. 207-209, K. Tjebbes and Chr. Wriedt call attention to the ocourrence of a dominant black in cats and suggest that it is the key to the much-discussed problem of the tortoiseshell male.
Ordinary black in cats is recessive to the ticking which gives tabby. Tjebbes (1924) crossed a tabby male with a Siamese female and obtained all blacks in the F1 generation. These blacks crossed inter se gave, amongst others, two tabby kittens. The black of the F1 generation was, therefore, obviously dominant to tabby. Tjebbes and Wriedt now give the results of three additional matings of the same type, and these confirm the occurrence of this dominant black.
They also record the mating of a Siamese female with a "striped yellow male” and say that three tortoiseshell kittens were produced, two males and one female. On the strength of this result they suggest that dominant black is probably the explanation of all tortoiseshell males.
The tortoiseshell male is of outstanding interest in regard to the problem of the inheritance of coat-colonr in cats and also in connection with the larger question of the chromosome theory both of inheritance and of sex-determination. It is, therefore, especially desirable that any theory of his origin should be examined with the greatest care. When so examined, the suggestion of Tjebbes and Wriedt does not seem to be in harmony with the facts of the case.
From Tjebbes’ own experiments it is clear that his dominant black is not affected by the factor for ticking, which gives tabby. If tortoiseshell males were the result of the combination of yellow and this dominant black, it should be impossible ever to get one with his black changed to tabby, yet such a tabby-tortoiseshell male is at present in our possession. His coat is a mixture of yellow and tabby arranged in irregular patches, with white chest and feet. Such tabby-tortoiseshells are common amongst females, but males are extremely rare. Fanciers refer to them casually, but only one other, to our knowledge, has been definitely recorded. It is practically oertain that the only difference between a black and a tabby lies in the presence of a factor for ticking (Doncaster, 1913, p. 21 footnote; Whiting, 1918 and 1919; Bamber, 1927, pp. 5-13), so that our male is a tortoiseshell with the ticking factor added.
It is, of course, possible that two doses of ticking might give tabby with one of dominant black although one dose, as in Tjebbes’ results, has no visible effect, but it is unlikely that the "tabby” so produced would be indistinguishable from normal tabby. The fact that the large patches of tabby on our male are perfectly normal suggests that dominant black is not the key to his exceptional colouration. He is fertile but has been in our possession for a short time only and so far his offspring[1] throw no light on the present subject.
[1. These breeding results have not yet been published, They will appear later when more facts have been collected.]
Another point in Tjebbes’ and Wriedt’s paper calls for comment, They record two tortoiseshell males and one tortoiseshell female from the cross yellow male x Siamese female. But a yellow male does not normally transmit yellow to his sons — yellow being apparently sex-linked (Little, 1912 and 1919; Doncaster, 1912 and 1913, p. 19; Bamber, 1927, p. 43; Bamber and Herdman, 1927, pp. 88-94). Here he appears to have done so in two cases in one litter. In view of the extreme difficulty of sexing some kittens at birth, it will not be considered unduly critical if we express a hope that these recorded tortoiseshell males will either be allowed to grow up, or be dissected. Some kittens are quite unmistakably male, others as unmistakably female, but some are so very indefinite that, even with kittens of both sexes in the same litter for comparison, it is not always possible to say to which sex they belong. There is therefore a possibility that if Tjebbes’ and Wriedt’s kittens are still young, they may have been sexed incorrectly. We dare to suggest this only because the occurrence of two males in one litter both inheriting yellow from their father is such a very extraordinary result.
Taking for granted, however, that the sexes are correctly recorded, the occurrence of the two tortoiseshell males is no justification whatever for the assumption that dominant black is the key to the problem. These males got their yellow from their father and that is abnormal. As Tjebbes and Wriedt seem to suggest, it may have been due to crossing-over between the X- and the Y-chromosome in the yellow male — yellow being sex-linked. But that alone would be sufficient to explain the results, and the dominance of the black is superfluous (Doncaster, 1912 and 1913, pp. 21 and 22; Bamber, 1927, pp. 19-23). If they mean to insist on the dominance of the black, as they evidently do, this implies that they have overlooked the fact that it is the presence of the yellow in their male kittens which is unexpected, not the presence of the black.
It will be interesting to see the results of further matings involving dominant black and yellow.
REFERENCES.
Bamber, R. C. (1927). Bibliographia Genetica, in, pp. 5-52.
Bamber, R. C. and Herdman, E. C. (1927). "The inheritance of black, yellow and tortoiseshell coat-colour in cats.” Journ. Genetics, xviii, no. 1, pp. 87-97.
Doncaster, L. (1912). "Sex-limited inheritance in cats.” Science, N.S., xxxvi, p. 144.
Doncaster, L. (1913). "On sex-limited inheritance in cats and its bearing on the sex-limited transmission of certain human abnormalities.” Journ. Genetics, in, no. I, pp. 15-22.
Little, C. C. (1912). "Preliminary note on the occurrence of a sex-limited character in cats.” Science, N.S., xxxv, pp. 784-785.
Little, C. C. (1919). "Colour inheritance in cats with special reference to the colours black, yellow and tortoiseshell.” Journ. Genetics, viii, no. 4, p. 281.
Tjebbes, K. (1924). "Crosses with Siamese Cats.” Journ. Genetics, xiv, no. 3, pp. 365-366.
Tjebbes, K. and Wriedt, Chr. (1926). "Dominant Black in Cats and its bearing on the Question of the Tortoiseshell Males.” Journ. Generics, xvn, no. 2. pp. 207-209.
Whiting, P. W. (1918). "Inheritance of coat-colour in cats.” Journ. of Exp. Zool., xxv, pp. 540 and 550-559.
Whiting, P. W. (1919). "Inheritance of white-spotting and other colour charaeters in cats.” Amer. Naturalist, Lin, pp. 474-477.
DOMINANT BLACK IN CATS AND TORTO1SESHELL MALES. A REPLY. By K. Tjebbes and Chr. Wreidt.
Journal of Genetics, 1927.
The criticism by Mrs R. C. Bisbee and Miss E. C. Hardman in the June number of this Journal (Vol. XVIII. p. 219) was rather welcome to us, since, in our reply to it, it gives us an opportunity of publishing some more details on our tortoiseshell males. Before doing so, however, we wish to say that we do not regard our suggestion that the occurrence of dominant black may have something to do with the occurrence of tortoiseshell males as a final interpretation for all such males. But it seems to us that our case of two tortoiseshell males in a single litter of three tortoiseshells from a cross with dominant black was distinctly suggestive, and that there is a greater possibility that the few tortoiseshell males mentioned in the literature are due to the occurrence of dominant black than to either non-disjunction or freemartinism.
It is beyond all doubt that dominant black occurs in our Siamese, material; it is also beyond doubt that this black does not lie in the sex-chromosome. There is, of course, a possibility that there are Y-chromosomes containing a factor which, in co-operation with dominant black, causes tortoiseshell. But we are quite convinced that the question of the tortoiseshell males is still unsolved, and that many experiments must yet be carried out before it will be possible to solve it.
With regard to our two tortoiseshell males already mentioned, we regret to state that one of them died when only two days old. He was however dissected and proved to be an indubitable male. The other one is still alive. He is now 17 months old, but so far has produced no off¬spring, although several queens were offered to him. Most probably he has never copulated, and we are forced to suppose that he is abnormal, or at any rate infertile. As the mother of these cats is still alive and vigorous, we are now trying, among other experiments, to repeat the cross that gave such an astonishing result.
TORTOISESHELL TOMCATS AND FREEMARTINS: A NOTE ON THE OCCURRENCE OF FUSED PLACENTAE IN CATS - T. Hume Bissonnette
Journal of Heredity, Volume 19, Issue 2, 1 February 1928, Pages 87–89
In 1920 Doncaster suggested that the sterility of the rare Tortoiseshell Tomcats may be due to sex-reversal by the action of a male sex-hormone during foetal life in the same way as in the freemartin in cattle, as shown so conclusively by Lillie, Bascom, Bissonnette, Keller and Tandler, and others. He suggested that the reversal might be carried a step farther and produce morphological but non-functional males, the sterile Tortoiseshell Toms. In that year he and Bamber began a search for fused placentae in cats, with the vascular anastomosis between the placental circulations necessary to make this possible. After his death this search was continued by Bamber who reported in 1922 on the negative results of the investigation. Among some 253 cat placentae examined she found some placentae coherent in the uterus, yet she was of the opinion that no confluence of blood vessels from the two embryos occurred in her specimens. The thin membranous parts of the chorions wero coherent but the highly vascular zonary placentae proper, with villi, were not fused completely enough to give the vascular connection necessary to produce this type of sex-reversal. She states, however, that she sees no reason to doubt the possibility of such union as she found being carried a step farther and producing the confluencc required or postulated by Doncaster. She thinks, however, that the sterility is conditioned by genctic or chromosomal abnormality and in a later paper she and Herdman offer evidence in support of the genetic basis for the occurrence of these sterile males but criticize the explanation offered by Tjebbes and Wriedt along somewhat similar lines.
Little discussed the suggestion of Doncaster and discarded it in favor of a genetic explanation for the sterile males. He suggested non-disjunction of the sex-chromosome rather than hormonic sex-reversal.
Hughes has shown that in swine the fusion of placentae is accompanied in rare cases by the same type of intersexuality as is found in the freemartin of cattle. Hoadley with a sin-gle case, demonstrated at the meetings of the American Association ior the Advancement of Science in Philadelphia December, 1926. supports Hughes’ findings. These cases show beyond question that it is not alone among cattle that such a mechanism for the transfer of sex-hormone from embryo to embryo in utero, is to be found. as was supposed by many.
Hartman mentioned a case among dogs, reported to him in a private communication by Professor H. M. Evans of California, in which two very young embryos were found in an apparently single placcnta, which he considered to have resulted from fusion of two originally discrete placentae crowded together in the uterus. ln this paper Hartman also refers to some other cases reported long ago, before the true nature of the freemartin was recognized, in which some abnormalities in man and the dog were classed as of the freemartin nature.
Bonnevie reported on a sterile fe male tortoiseshell cat which resembled the freemartin in some respects, but she accounts for the condition on the basis of chromosomal disturbances. She cites Oshima and Oyama as also account [. . .]
A CASE OF POTENTIAL FREEMARTINS IN CATS - Thomas Hume Bissonnette, Trinity College, Hartford, Connecticut
The Anatomic Record. Vol. 40, No. 3
In a preliminary paper the author discussed the genetic significance of fused placentae in cats (1928c). The findings of Hughes and Hoadley for swine, of Lillie, Keller, Bisson* nette, and others for cattle, and of Bonnevie for cats, were also discussed as bearing on the wider application of the theory of tlie freemartin of Lillie and Keller and Tandler. It was shown that Doncaster (1920) and Little (1920) were within the bounds of possibility whcn they suggested that placental vascular anastomosis between male and female cat embryos might be the cause of the sterility of male tortoiseshell cats, since the case here described is evidence that such anastomosis does sometimes occur and at a stage in development preceding visible sexual dilTerentiation, just as in cattle. Lack of such a case as the one described in the present article caused Bamber (1922) to seek to account for the sterile tortoise-shell tomcats ön a genetic basis.
MATERIAL AND METHODS
This pair of twin kittens, members of a litter of seven, was found in a cat dissected bv the class in comparative vertebrate anatomy in Trinity College. The cat had been killed the previous summer with illuminating gas, injected through the aortic arch with a carmine-starch-formalin injection mass for arterial dissection, and then skinned before being immersed in 10 per cent formalin for storage. At the time of injection, the abdomen had been partly opened to admit the preservative.[. . .]
THE INCIDENCE OF STERILITY AMONGST TORTOISESHELL MALE CATS
Ruth C Bamber (Mrs Bisbee) and E Catherine Herdman.
Journal of Genetics , Volume 24, Issue 3 , pp 355-357, 1931
Tortoiseshell [1. Tortoiseshell here includes tortoiseshell-and- white and tabby-tortoiseshell] coat-colour in cats is the heterozygous condition of yellow and black. Tortoiseshell females are fairly common, but in the ordinary course of inheritance tortoiseshell males do not occur. Apparently both black and yellow are sex-linked and so do not normally occur together in the male (Little, 1912 and 1.919; Doncaster, 1913; Bamber, 1927 (survey of literature); Bamber and Herdman, 1927 a). Tortoiseshell males do appear, however, as rare exceptions, and it is generally stated in scientific literature that such males are usually sterile.
Since considerable importance is attached to this sterility in practically all discussions on the origin and gametic constitution of the abnormal colour type, it is of interest to ascertain exactly what data exist on the subject. The recorded facts are few. Cutler and Doncaster, in 1915, gave a list of the then known adult tortoiseshell males with the available information regarding their fertility. They recorded seven in all. Of these one had died without being mated. Of the remaining six, one was certainly fertile and two were certainly sterile. Another was certainly almost sterile [2 He mated repeatedly without result, but on one occasion a female, after mating with him, produced one kitten. As, however, she had not been kept in confinement after the recorded mating, the paternity of the kitten is uncertain.]. The remaining two were both said to have sired kittens, but there was no definite proof.
Since this publication, seven other adult tortoiseshell males have come under our notice. One of these, “Lucifer,” has been recorded by us elsewhere (Bamber and Herdman, 1927, 1928). The others are not on record in scientific liteJature, but some of them are registered in the books of the Cat Fancy.
Table I contains all the adult tortoiseshell males hitherto recorded in scientific literature, together with those now recorded for the first time. In order to avoid accidental duplication of records we have given the name of each cat and the name of his owner. We have been able to identify the cats recorded by Cutler and Doncaster by reference to Doncaster’s original correspondence. Eor most of the new records we are indebted to Sir Claud Alexander.
TABLE I.
Thus, of the fourteen adult tortoiseshell males now on record, three died unmated and one is savage and will not mate. Of the remaining ten, two are reputed to have been fertile but there is no recorded evidence. There are only eight, therefore, of which we have definite [2. In all these cases matings have been made under definitely controlled conditions, the females having been kept in confinement both before and after mating] knowledge in regard to fertility. Of these, three are certainly fertile, four are certainly sterile and one is certainly almost sterile, probably completely so.
There are no published records of the incidence of sterility among ordinary male cats, but in the course of our own breeding experiments we have used fourteen such males, taken at random from the general population, and all have been fertile. It does appear, therefore, that the abnormal association of black and yellow in the male cat is correlated with a tendency towards sterility.
Our thanks are due to the Royal Society of London for grants which enable us to investigate the genetics of domestic cats.
REFERENCES.
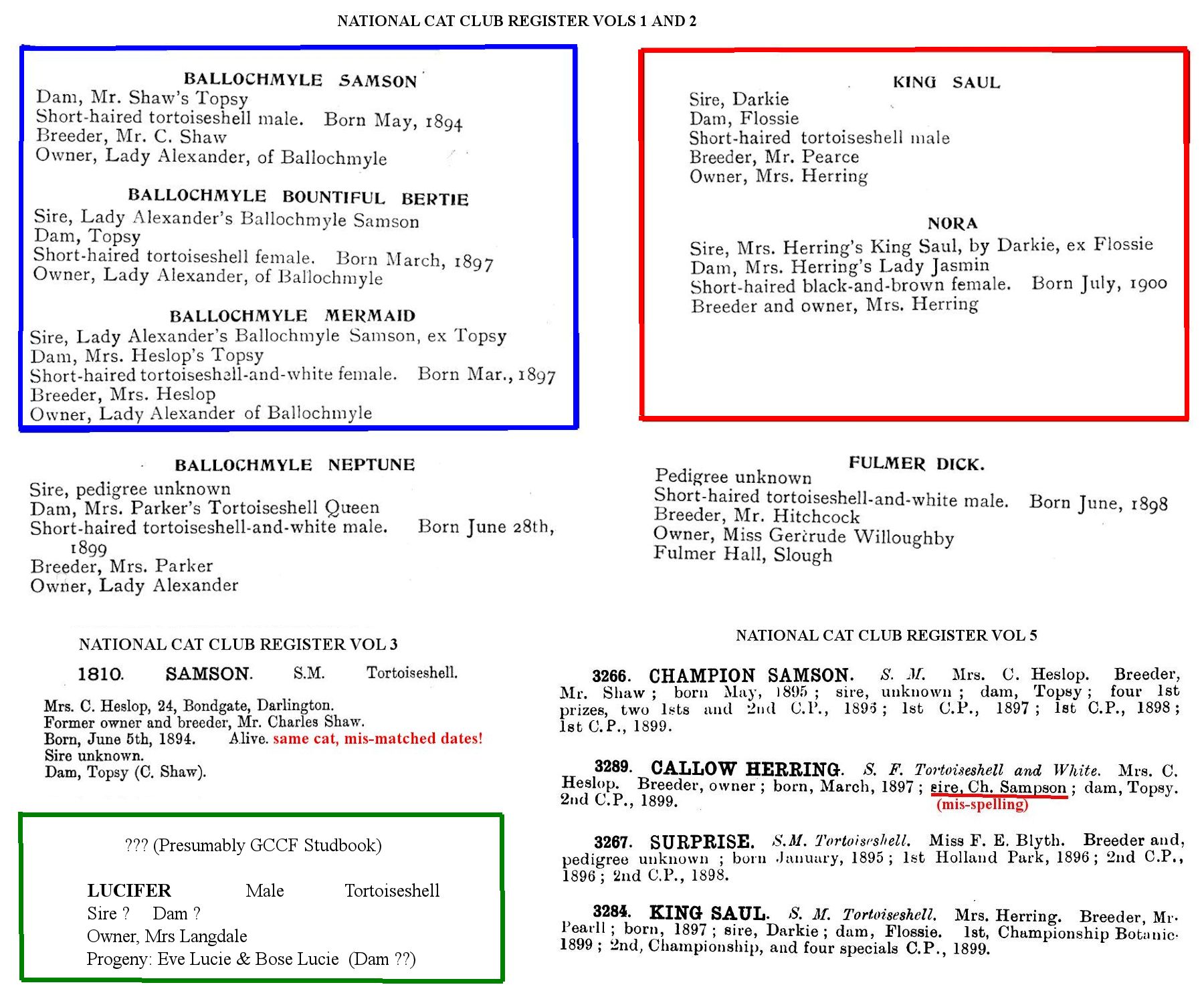
A REPORT ON THE PROGENY OF A TORTOISESHELL MALE CAT, TOGETHER WITH A DISCUSSION OF HIS GAMETIC CONSTITUTION.
Ruth C Bamber (Mrs Bisbee) and E Catherine Herdman.
Journal of Genetics , Volume 26, Issue 1 , pp 115-128, 1932
In the normal course of inheritance in cats tortoiseshell males do not occur.
(Doncaster, 1904, 1913; Little, 1912; Whiting, 1915, 1918; Bamber and Herdman, 1927 a.)
Thus tortoiseshell is the heterozygous condition of black and yellow, but appears normally in the female only.
In addition to the normal breeding results certain exceptions occur from time to time. Thus sometimes a yellow male sires a black daughter, and very occasionally a tortoiseshell male appears. There is also one case on record of a yellow female having bred as a tortoiseshell, and another case where the cross yellow male x yellow female gave three tortoiseshell females. There is also one record of an anomalous yellow female from the cross yellow male x tortoiseshell female. This anomalous yellow female was apparently homozygous for yellow, yet had a small black spot which she transmitted, together with the yellow, to a son. All these exceptions are rare and many, theories have been put forward to account for their occurrence. (For a survey of the literature on these exceptional individuals see Bamber, 1927, pp. 18-44. See also Bamber and Herdman, 1927a.)
Tortoiseshell males, when they do appear, are in many cases sterile. Three only are recorded as certainly fertile (Bamber and Herdman, 1931).
The first of these, Samson, was owned by Sir Claud Alexander, and was used by him for breeding under controlled conditions. Since, however, the matings were not made for the purpose of definite scientific investigation, the records of the progeny were not sufficiently complete to reveal his gametic constitution (Doncaster, 1904, 1913).
The second fertile tortoiseshell male, unnamed, is also the property of Sir Claud Alexander. So far only one experimental mating with this cat has been recorded (Bamber and Herdman, 1931).
The third fertile tortoiseshell male has been in our own possession since 1926 and has been recorded by us elsewhere (Bamber and Herdman, 1927 b and c; 1931). He is the only male of this type which has hitherto been available for purely scientific breeding experiments. Unfortunately he has recently died; therefore, although our experiments are not completed, there is no reason for further delaying the publication of our results.
Before this cat came into our possession he was owned by Mrs Langdale and was registered by her in the books of the Cat Fancy as “Lucifer.” He had already sired two kittens in a controlled mating, and these also were registered as “Eve Lucie” and “Bose Lucie.” Apart from these all his known progeny have been sired, in our own experimental matings.
In colour Lucifer was tabby-tortoiseshell and white. Tabby-tortoiseshell is tabby and yellow, whereas ordinary tortoiseshell is black and yellow. Tabby is black with a ticking factor added. This ticking factor has no visible effect on the yellow, and its inheritance is quite inde-pendent of both black and yellow. Its presence, therefore, does not in any way complicate our breeding results: for the present purpose it may be ignored, tabby being counted as black. The white may also be disregarded, white spotting being due to a separate factor or factors whose inheritance is quite independent of that of black and yellow.
The matings herein recorded have been made under absolute control. The females were kept in entirely closed pens and were never allowed out. The only times they met a male were when they were deliberately mated. All the crosses have been made in our own cattery, except No. XIV (Table I), and that was made under conditions exactly similar to our own. The parentage of the kittens herein recorded is, therefore, absolutely certain.
Lucifer — male (14) — has been mated to black, yellow and tortoiseshell females.
* Anomalous yellow is yellow with a minute amount of black spotting. Its gametic constitution is discussed below.
Fuller details of these crosses are given in Table I.
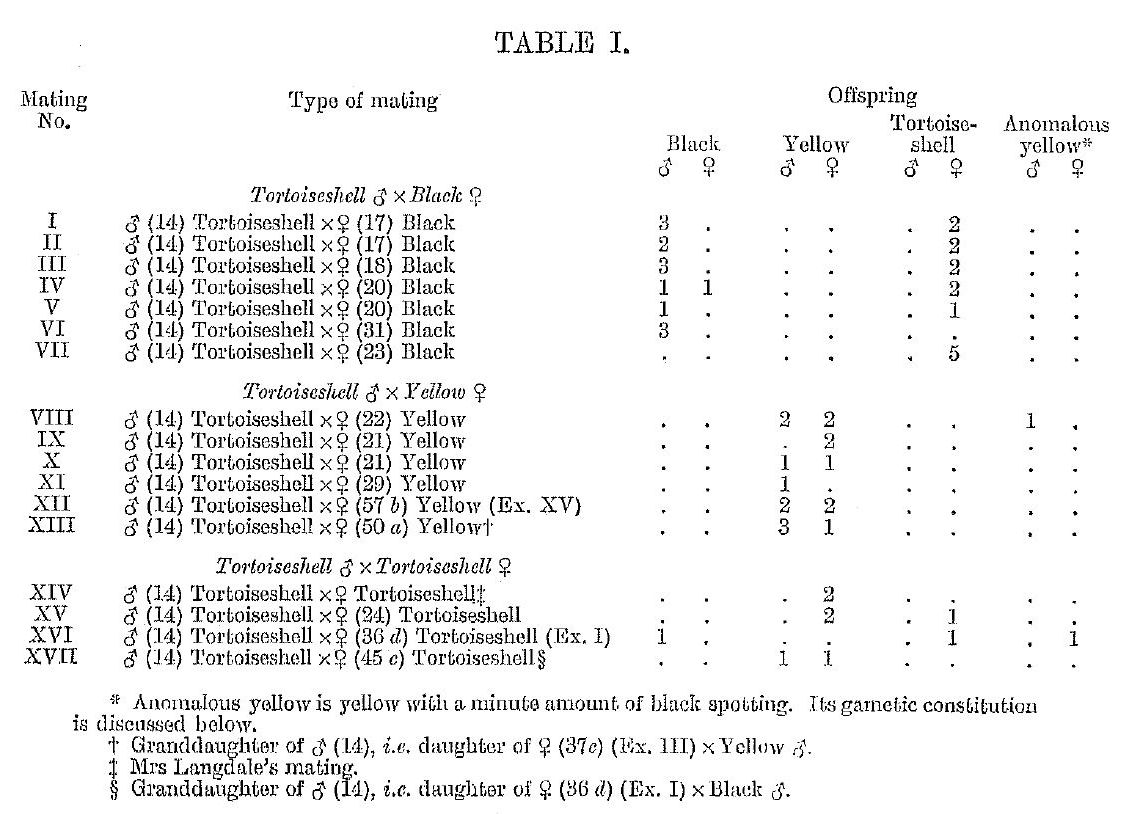
Comparison of these results with the normal breeding results and known exceptions, summarised above, reveals the fact that, apart from the production of the one anomalous yellow male (Mating VIII, Table I), this tortoiseshell male (14) has bred exactly like a yellow male. This is equally true whether he was mated to related or unrelated females (cf. Table I). Moreover his daughters, mated to unrelated males, gave entirely normal results; also one mating between a grandson and a granddaughter gave a perfectly normal result. Details of these matings are given in Table II.

Unfortunately we are not able to give any data in regard to the offspring of his sons. A few were chloroformed when newly born and some died before reaching maturity. On dissection most of these proved to be quite normal. One newly born kitten, however, had one testis abdominal and another had both testes abdominal, but undescended testes in a newly born kitten may have no special significance. Ten sons of male (14) grew up and one only is known to have been abnormal. This animal, a yellow male (48a) from mating X (Table I), lived to be 2 years old and was externally a male but his testes were not descended and he never showed the slightest sex instinct. Other tom cats treated him simply as a kitten — they did not resent his presence even when females were in season. He was eventually chloroformed and on dissection his testes were found to be represented by two rounded bodies, very unlike testes in appearance, connected to the vasa deferentia and situated in the abdominal cavity. This cat will be described in detail in a separate publication when his gonads and other endrocrine glands have been fully investigated histologically. Cats with undescended testes do sometimes occur in the general population. Whether or not they have any connection with the phenomenon of the tortoiseshell male is not known.
Discussion.
It appears to us that the facts herein recorded throw some light on the much discussed problem of the origin and gametic constitution of the tortoiseshell male cat. At any rate they disprove many of the theories which have been put forward, and so clear the ground for discussion.
The problem of the exceptional tortoiseshell male is, of course, inseparable from that of the normal inheritance of black, yellow and tortoiseshell. The ordinary facts of inheritance can be explained equally well by assuming:
(1) That one colour only is sex-linked and is completely dominant to the other in the male, incompletely dominant in the female, i.e.
(a) that yellow is sex-linked and that black is present in all gametes, one dose of yellow being completely dominant to all black in the male, incompletely dominant in the female (Doncaster, 1912 and 1921);
or (b) that black is sex-linked and that yellow is present in all gametes, one dose of black being completely dominant to all yellow in the male, incompletely dominant in the female.
(2) That both black and yellow are sex-linked:
(a) as a pair of allelomorphs (Little, 1912; Doncaster, 1913; Whiting, 1915 and 1918; Ibsen, 1916);
or (b) with yello w epistatic to black, black being present on all X-chromosomes (Little, 1919);
or (c) with black epistatic to yellow, yellow being present on all X-chromosomes.
(For a survey of the literature and discussion of the above theories see Bamber, 1927, pp. 14-17 and p. 43.)
In view of the fact that all yellow cats examined by us, both males and females, show a few black (or tabby) hairs it seems improbable that there is any difference in dominance in the two sexes (Bamber and Herdman, 1927 a). If there is not a sex difference in dominance the only alternative seems to be that both yellow and black are sex-linked. The fact that yellows do occasionally give black in their offspring, whereas blacks are not known to give yellow, makes it unlikely that yellow is hypostatic to black. There remain, therefore, two hypotheses in regard to the normal inheritance of black and yellow which seem to us more free from objection than the rest, namely that black and yellow are a pair of sex-linked allelomorphs, or that black is present in all X-chromosomes and is hypostatic to yellow, yellow and its allelomorph (which permits the development of black) being also sex-linked.
To account for the tortoiseshell male many theories have been put forward and, naturally, each one rests upon the particular scheme of normal inheritance accepted by its author. Even in the light of the facts of normal inheritance some of these earlier theories break down, but many remain as possibilities. All the theories put forward before 1924 have already been critically surveyed by one of us elsewhere (Bamber, 1927, pp. 18-44) and therefore will not be discussed at length in the present paper. Reference will be made to those alone which rest upon a scheme of normal inheritance which is not impossible in the light of the known facts. Also they will be- discussed only in relationship to the new facts herein recorded.
Crossing-over, mutation, extension factors, sex-reversal, non-disjunction and dominant black have all been suggested as the cause of the occasional tortoiseshell males.
Crossing-over in the male appears to be the key to the suggestions made by Doncaster (1912, 1913) to account for the occurrence of the tortoiseshell male [1. Crossing-over in the male is always quoted as Doncaster's theory but actually he did not use the term—he said “failure of sex-linkage.”]. In 1912 he believed black to be present in all gametes and yellow to be sex-linked, one dose of yellow being completely dominant to all black in the male, incompletely dominant in the female. He suggested that if, owing to failure of sex-linkage, a yellow male gave yellow to a son the yellow so received might not act normally in regard to black, and so might give a tortoiseshell male instead of a yellow. In 1913, accepting the possibility that both black and yellow are sex-linked, he suggested that, owing to failure of sex-linkage, a male might give his own colour to a son, and this, if it met the alternate colour received from the mother, would make the son heterozygous, and therefore tortoiseshell. Hayes, in 1923, made a similar suggestion.
If crossing-over in the father were the cause of a tortoiseshell male he should carry either black or yellow in his 0-chromosome and should therefore give tortoiseshell sons either when mated to yellow females or when mated to blacks, But tortoiseshell male (14) gave yellow sons, with one exception, when mated to yellow females, and black sons when mated to blacks. Clearly his 0-chromosome was not carrying colour, and he could not, therefore, have been the result of crossing-over in his sire.
Crossing-over in the female was suggested by Ibsen (1916) as the cause of the exceptional tortoiseshell males. He accepted both black (B) and yellow (b) as sex-linked, but postulated also an additional factor for tortoiseshell (T), usually linked with yellow and unable to act except in the presence of black. He suggested that crossing-over in a tortoiseshell female, bTBt, giving gametes BT and bt, could give a tortoiseshell son of the constitution BT"-".
A tortoiseshell male arising in this way should give tortoiseshell daughters both when mated to black females, BtBt, and when mated to yellows, bTbT. This, of course, male (14) did not do. Ibsen suggests that a daughter of the constitution BTBt would be tortoiseshell, but that one of the constitution BTbT might be yellow owing to extreme restriction of black by the double dose of T to one dose of B. Granting these somewhat remote possibilities a tortoiseshell male of the constitution BT"-" might breed as a yellow, but even so, as Ibsen points out, all Iris daughters should give tortoiseshell to half their male offspring no matter to what males they be mated. We have unfortunately obtained only four sons, to date, from Lucifer’s daughters, so that the absence of tortoiseshell males is not conclusive. It is clear, however, that no daughter of a tortoiseshell male should, on Ibsen’s hypothesis, produce both yellow and black sons — each one should produce black sons and tortoiseshell sons, or yellow sons and tortoiseshell sons. Lucifer’s daughter (36 d) produced a black son in mating XVI (Table I) and a yellow son in mating XVIII (Table II). Evidently neither of her X-chromosomes was of the constitution BT, Lucifer, therefore, does not appear to have been the result of crossing-over in his mother.
Crossing-over, either in the male or the female, fails, therefore, to account for the origin of our tortoiseshell male in the light of his breeding results.
Mutation was suggested by Little in 1912 as the cause of tortoiseshell males. He supposed black and yellow to be sex-linked allelomorphs and suggested that tortoiseshell may arise as a separate colour factor by mutation. Thus a tortoiseshell male would not be heterozygous for black and yellow — he would simply carry a factor for tortoiseshell.
Such a male would not be expected to give either yellow or black daughters, but only tortoiseshells. And no daughter, whatever her colour, should be able to produce both yellow and black sons. She should produce black sons and tortoiseshell sons, or yellows and tortoiseshells, as according to Ibsen’s theory discussed above. But Lucifer produced yellow daughters and tortoiseshell daughters, and one of these latter (36 cl) produced a black son and a yellow son.
Mutation, therefore, will not account for tortoiseshell male (14) in the light of his breeding results.
Extension factors were suggested by Whiting in 1915 and 1918 as the clue to the problem of the tortoiseshell male. In the form in which he stated his theory it could not possibly stand, even apart from our own results reported above. He accepted black and yellow as sex-linked allelomorphs and suggested that a yellow male may become tortoiseshell by an extreme condition of black extension factors. If black and yellow are sex-linked allelomorphs a yellow has no black for the extension factors to act upon. If, however, black is hypostatic to yellow (or vice versa) the possibility of the action of extension factors does arise.
If such factors are present they should be transmitted and should give some recognisable effect in the offspring. There is little or no evidence of the action of such factors in Lucifer’s progeny. True, he sired one unexpected black female (mating IV, Table I), but this is an exception which has been recorded comparatively often from yellow males mated to ordinary females with no suggestion of special modifying factors. There are also the two anomalous yellows - yellow with a minute amount of black spotting — amongst Lucifer’s offspring, and these might well be claimed as evidence of the presence of modifying factors. Both died as kittens, so there is no evidence as to their gametic constitution, and theoretically they could have been either yellows with black extension factors or blacks with yellow extension factors. Since Lucifer bred almost entirely as a yellow, yet was a perfectly ordinary tortoiseshell in appearance, he must have been a yellow with extremely strong black extension factors, if extension factors were involved at all. Yet in all his 56 offspring there were only 3 in which any sign of such extension factors could be claimed. The remaining 53 were perfectly normal in colour, his tortoiseshell daughters being ordinary medium tortoiseshells. Even discarding his 14 black offspring, where black extension factors would give no visible effect, there still remain 42 individuals which should show the effect of black extension factors if they were present, and, of these, only the 3 referred to above could possibly be claimed as giving any sign of such factors.
It seems to us extremely improbable, therefore, that extension factors were responsible for the tortoiseshell colouration of: male (14).
Sex-reversal, from female to male, was suggested by Wright in 1918 and. by Doncaster in 1920 as the cause of tortoiseshell males. Thus a tortoiseshell male would be XX in constitution, and would be heterozygous for black and yellow, supposing both to be sex-linked. Such an XX male should give only female offspring, whereas male (14) gave both males and females. Sex-reversal from female to male, therefore, does not account for tortoiseshell male (14) in the light of his breeding results.
Slight sex-reversal from the male to the female condition was suggested by Bamber in 1922 as the cause of the tortoiseshell colouration of exceptional males. If yellow alone is sex-linked and black is present in all gametes, there being a difference in the degree of dominance of yellow in the sexes, then, a slight change from the male towards the female condition (as in some of Goldschmidt’s moths) might be sufficient to allow the female colouration to appear.
Such, a male should breed as a yellow, and this is what male (14) did, apart from the production of the one anomalous yellow male. And even this exception might be looked upon as another example of slight sex-reversal. There is nothing in the breeding results of male (14) to prove that he was not an example of slight sex-reversal.
Non-disjunction through three generations was used by Little in 1919 in a complicated theory of the origin of tortoiseshell males. He believed black, B, to be present in all X-chromosomes and yellow to be due to a factor, Y, for the restriction of black from the coat, its allelomorph, y, allowing the extension, of black to the coat. Y and y he believed to be sex-linked, one dose of Y being normally completely epistatic to one dose of B. Thus:
Yellow male = YBX0;
Black male = yBX0;
Yellow female = YBXYBX;
Black female = yBXyBX;
Tortoiseshell female = YBXyBX.
He pointed out that from a cross between a non-disjunctional tortoiseshell female and a yellow male an individual "-"YBX could occur, receiving YBX from the father and - from the mother [1. As Little pointed out a similar individual could arise from a non-disjunctional black female mated to a yellow male. Actually a non-disjunctional female of any colour could give the same result]. He suggested that the absence of the 0-chromosome might “upset the somatic relation of certain of the characters whose factors are carried by the X-chromosome” and so "-"YBX might give tortoiseshell instead of yellow. He suggested that the absence of the 0-chromosome might also make such an individual sterile, as are so many tortoiseshell males. From the same mating females of the constitution YBXyBX0 could arise, and such a female, if mated to a normal yellow male, YBX0, could give, amongst others, an individual YBX00, which Little calls a peculiar yellow male. He suggests that if such a male were mated with any female showing primary non-disjunction individuals of the constitution YBX and "-"YBX0 could be produced receiving respectively YBX and YBX0 from the male. The "-"YBX individual, as in the first cross (see above), might be tortoiseshell and sterile, and Little suggests that the "-"YBX0 individual might be a fertile tortoiseshell male — tortoiseshell because the 0 received from the father together with X might not exert a full normal influence until gametogenesis, yet fertile because of the presence of the 0 at gametogenesis.
According to Little such a fertile tortoiseshell male would be expected to breed as a yellow, and this is what male (14) did, apart from the production of the anomalous yellow male. Presumably an even further supposition could be made to account for him. A 0-chromosome having gone through as many vicissitudes as that of Little’s theoretical, fertile tortoiseshell male might also be excused for not exerting a quite full influence in an occasional son. In the light of the breeding results of male (14) Little’s hypothesis, therefore, still remains as a theoretical possibility.
Dominant black was suggested by Tjebbes and Wriedt in 1926 as the cause of some tortoiseshell males. In a cross Siamese female x Yellow male they obtained two tortoiseshell males and suggested that the dominant black of Siamese may not behave like the recessive black of ordinary cats in regard to yellow. Very little is known about dominant black, but Tjebbes and Wriedt (1927) state that it is not sex-linked. That being true a tortoiseshell male should give this black to half his offspring of both sexes, and in his sons this should give tortoiseshell as in his own case. Lucifer produced ten yellow sons, and one with a minute amount of black in no way comparable with himself. He produced no tortoiseshell sons. It is very unlikely, therefore, that he was carrying dominant black.
From the breeding results of our own tortoiseshell male, therefore, it is almost certain that he was not the result of the combination of dominant black and yellow.
Thus from the results herein recorded it is quite certain that neither crossing-over in the male, nor crossing-over in the female, nor mutation, nor sex-reversal from female to male could possibly account for tortoiseshell male (14). It is also practically certain that neither extension factors nor dominant black can account for him.
The two theories which do not completely fail in the light of these new facts are the theory of slight sex-reversal towards the female condition and Little’s theory of non-disjunction through three generations. And both these hypotheses are open to serious objections.
In regard to the possibility of slight sex-reversal in the female direction it should be noted that male (14) was an exceptionally large cat with a very broad masculine head and that his sex instincts were extremely well developed. There was not the slightest suggestion of femaleness about him apart from his colour. Further, sex-reversal of this type could only be expected to affect the colour if there were normally a sex difference in the degree of dominance of yellow over black, or of black over yellow. Facts reported by us in 1927 a make it very improbable that such a sex difference exists (see above).
In regard to Little’s hypothesis there are also difficulties. In the course of the complicated process which, he suggests, individuals of abnormal gametic constitution would occur at every step (cf. the details of Little’s hypothetical crosses given in the Appendix), and for the existence of such individuals there is no evidence. Also Little’s suggestions in regard to colour changes are not easy to justify. There seems no reason to suppose that in the male "-"YBX the YBX would give tortoiseshell instead, of yellow because of the absence of a 0-chromosome - in the female, YBXYBX give yellow and there is no 0 present. Also it is difficult to imagine why, when a male receives YBX and 0 together from the male, the YBX should give tortoiseshell instead of yellow. Moreover, the fact that male (48 a), though completely sterile and undeveloped sexually, was nevertheless yellow, not tortoiseshell, seems to us to argue against Little’s supposition that the colour reactions of black and yellow would be upset by sexual abnormality. Nevertheless Little’s hypothesis does remain as a theoretical possibility.
It seems to us, however, that it is possible to explain all the known facts in a comparatively simple way. We suggested in 1927a that partial non-disjunction may sometimes occur in the female, so that a part of one X-chromosome remains attached to the other. This suggestion was made to account for a minute amount of black spotting which appeared in a yellow female that proved, on breeding to be homozygous for yellow. She arose from a cross tortoiseshell female x yellow male, and she transmitted her minute amount of black together with yellow to one of her sons. Accepting the sex-linkage of both black and yellow it seems possible that this anomalous yellow female had received, from her tortoiseshell mother, a yellow-carrying X-chromosome to which adhered a small portion of the black-carrying X-chromosome bearing part of the factor for black. This suggestion, of course, involves the fractionation of the factor for black, and according to some workers a factor should be indivisible. Many facts, however, suggest fractionation (see Bamber and Herdman, 1927a and Bamber, 1929).
It seems to us that partial non-disjunction in the female would also best account for our own tortoiseshell male (14). We have never seen his mother, but his original owner who bred him stated that she was tortoiseshell. If, in the formation of the gametes of this tortoiseshell female, part of an X-chromosome, carrying black, failed to separate from the yellow-carrying X-chromosome, she might well produce a tortoiseshell son. He would be tortoiseshell because heterozygous for black and yellow, as is a female, but, if he received only a small part of the additional X-chromosome, it might not be sufficient to modify his sex. Such a male in forming his gametes might well tend to throw out the odd bit of chromosome. (The throwing out of a chromosome or part of a chromosome is not an unknown phenomenon.) If the elimination of this fragment were complete he would breed as a yellow, which is what male (14) did, with one exception. If, on the other hand, the elimination were not always quite successful, he might give some black to a few of his offspring, and this could account for the one exception, the anomalous yellow male [1. It would equally account for the anomalous yellow female, but as her mother was tortoiseshell the origin of her black is not certain.].
Conclusion.
We suggest that Lucifer, tortoiseshell male (14), arose by partial non-disjunction in his mother.
Our thanks are due to the Government Grant Committee of the Royal Society of London for grants which have enabled us to carry out the investigations reported in this paper.
APPENDIX.
Details of Little’s hypothetical crosses (1919).

REFERENCES.

The origin of the tortoiseshell male cat is an old puzzle of genetics. Bamber (1927) stated: "The normal mode of inheritance of black, yellow and tortoiseshell is only very imperfectly understood. - That black is sex-linked is widely accepted, but has never actually been proved. Either yellow or black or both are certainly sex-linked.” Komai in papers in 1952 and 1956, and Komai and Ishihara in 1956, had shown that the gene for orange (yellow) color was sex-linked, whereas the gene for black or tabby color was autosomal. Searle (1949) had made a similar census in London, and had reached the same conclusion. The effect of the gene for orange, in the homozygous or hemizygous state, was perfectly epistatic to that of the gene for black or tabby, and the orange color completely covered the black or tabby color. However, it is incompletely epistatic in the heterozygous state, resulting in the tortoiseshell coat.
[Hemizygous means a gene where only one copy is present instead of a pair. Genes on the X chromosome in male cats are expressed regardless of whether they are dominant or recessive because there is no corresponding gene on the Y chromosome.]
This was upheld by the results of censuses of cat coat colors and sexes (Searle 1949, Komai 1952, Suzuki 1953). In orange cats there were always many more males than females among these cats, but no such disparity in black or tabby cats. Darwin (1868) had stated, "The peculiar colour called tortoiseshell is very rarely seen in a male cat, the males of this variety being of a rusty tint." Doncaster (1904) had said "Orange females are very rare, although males are common." [orange females are less common than orange males, but are not rare]
Komai stated that gene which is allelic to the sex-linked gene for orange (O) is the one for non-orange or wild-type of orange (o+), and not the gene for black (b) or for tabby (b+) which is located on one of the autosomes. Komai found it striking that this plain fact had escaped notice of recent geneticists including Sprague and Stormont (1956) as that basic fact was essential for understanding the origin of tortoiseshell males.
Origin Of Tortoiseshell Males
Komai had previously stated that the mothers of all tortoiseshell male cats were tortoiseshells. Records of the births of 65 tortoiseshell males from various parts of Japan proved this untrue. They clearly showed that the mother of a tortoiseshell male could be of any color. That implied that she carried either O or o+ in her X-chromosome and either b or b+ in one of her autosomes. But, the sex-linked gene possessed by her and that possessed by her mate should be in a heterozygous relation 0/o+. Komai deduced that the tortoiseshell male was heterozygous for O and o+, much like the female tortoiseshell.
Tortoiseshell males should be hemizygotic, but heterozygous for O. He assumed that the Y-chromosome in such exceptional males carried O or o+ due to crossing-over between X and Y chromosomes in a germ cell of the father of the cat which would transfer the O or o+ from X to Y. By 1957, the X and Y chromosomes were known to be differentiated into pairing and non-differential segments, and chiasma [joins] could be formed between the pairing segments of the two chromosomes. Komai considered it likely that crossing over took place between X and Y. During that process, the sex-linked gene O or o+ might be transferred from X to Y and a resulting male offspring would get the gene in question. Komai considered this accounted for the occasional heterozygous male.
Regarding sterility in tortoiseshell males, Komai assumed the presence of a gene(s) governing male fertility was on the Y chromosome. If it transferred from X to X chromosome during crossing over then it would be absent from the male offspring getting that Y chromosome. He admitted that this might be revealed to be wrong by future studies. Komai found three specimens of fertile tortoiseshell males. Two were examined by Ishihara and their testes were perfectly normal and apparently fertile. No breeding data was been available for these cats. The third specimen had proven to be fertile. He assumed that the crossing-over between X and Y had failed to remove from Y the gene complex that governed male fertility, even though it did transfer O from Y to X. If so, a fertile tortoiseshell son should produce further tortoiseshell males. Scientific literature recorded three fertile tortoiseshell males. One had no breeding record. Another, reported by Cutler and Doncaster (1915, 1932) might have originated through Komai’s crossing-over hypothesis. The third was "Lucifer” (Bamber and Herdman, 1932) who was a genetic orange despite of its tortoiseshell appearance, but the covering effect of the orange gene had apparently become incomplete by the presence of a modifying gene or by some other cause.
It was difficult to present any cytological evidence for the crossing-over hypothesis because the sterile testes contained few mitotic or meiotic figures. Komai hoped that a tissue culture of somatic tissue would provide mitotic figures. In the 1950s it was hard to detect any structural change in the Y-chromosme. Komai hoped to identify and study tortoiseshell males born of the same mother, but with different fathers.
Of previous hypotheses, Komai noted that the freemartin hypothesis had been revived by Sprague and Stormont (1956). This postulated that female embryos could render male embryos intersex or infertile in utero [seen in cattle where a female twin causes feminisation in her male twin]. The gonads of sterile tortoiseshell males showed no sign of intersex/feminisation.
Komai concluded:
References included:
Hayes, F. A. 1923. The tortoiseshell cat. J. Hered. 14, 369-376.
Komai, T. 1946. On the inheritance of black, yellow and tortoiseshell colour in cats, and the problem of the tortoiseshell male. Proc. Jap. Acad. 22, 265-268.
Komai, T. 1946. On the inheritance of black, yellow and tortoiseshell colour in cats, with special reference to the problem of the tortoiseshell male. (in Japanese). Seibutu 1, 1-7.
Komai, T. 1947. A new hypothesis on the origin of tortoiseshell male cat. Mem. Coll. Sci. Kyoto Univ. ser. B, 19, 17-21.
Ohshima, H. and Oyama, J. 1925. On a tortoiseshell male cat with odd eyes. (in Japanese). Toyogakugei-zasshi 507, 9-18.
Searle, A. G. 1949. Gene frequencies in London's cats. J. Genet. 49, 214-220.
Tjebbes, K. and Wriedt, C. 1926. Dominant black in cats and its bearing on the question of the tortoiseshell males. J. genet. 17, 207-209.
Jude and Searle wrote that in many animals, sex was determined by a difference in one chromosome pair. The females having 2 similar X-chromosomes while the males one X- and one Y- chromosome. Whereas similar chromosomes paired along the whole of their length, the Y- pairs only partially with the X-chromosome. Sex-linked genes were those carried by the X-chromosome, e.g. the gene for yellow coat colour in the domestic cat. The yellow gene had a black counterpart (allelomorph) meaning that at a given point on an X-chromosome there may be found either the yellow coat colour gene or the black coat colour gene. A female could inherit one black and one yellow gene in which case it would be tortoiseshell colour.
According to these simple ideas of sex determination in mammals, it seemed that nature prohibited tortoiseshell males, for the male cat with its single X-chromosome could carry black or yellow, but not both. In defiance of theory, male tortoiseshell cats turn up on rare occasions, but are usually sterile. So far no one had been able to account satisfactorily for the occurrence or the sterility of those tomcats.
An even rarer phenomenon, the fertile tortoiseshell tomcat, was described by them in Nature magazine.
A FERTILE TORTOISESHELL TOMCAT – A.C. Jude and A.G. Searle 1957 (Nature)
Jude and Searle wrote that the gene for yellow coat-colour in the domestic cat was sex-linked; tortoiseshell cats were heterozygous for yellow and therefore normally female ; the rare tortoiseshell male is usually being sterile. However, a tortoiseshell male of known parentage had produced in the space of two years at least sixty-five offspring. 26 were recorded as female, 17 as male and 22 were unsexed. The kittens were the result of systematic matings carried out in confinement, so their paternity was not in doubt. The male concerned was "Blue Boy”, a dilute tortoiseshell English rex ; his father was ‘Kalli’, dilute yellow and the first English rex cat. His mother was ‘Serena’, a tortoiseshell heterozygous for rex and for dilute. Because the cats concerned were being used for establishing the rex breed, the results of matings were carefully recorded even before Blue Boy’s true nature was realized.
Eleven further tortoiseshell males were recorded among Blue Boy’s forty-three sexed offspring. Three of theoe males had reached maturity by 1957, but none had yet been proved fertile. Jude and Searle knew of no previous records of tortoiseshell males siring further tortoiseshell males ; one of the tortoiseshell males studied by Doncaster had produced tortoiseshell offspring, but their sex was apparently unrecorded.
Blue Boy was mated to 6 different tortoiseshell females, all related to him. The offspring included in each sex all three possible genotypes at the yellow locus ; that is, males and females which were phenotypically (i) black, blue or tabby, (ii) tortoiseshell or dilute tortoiseshell (blue-cream), (iii) yellow or cream. They thought most likely hypothesis was that in Blue Boy the yellow gene was only partially sex-linked instead of fully sex-linked. It would have to be present in the pairing segment of the Y-chromosome, instead of being absent from Y.
Using the notation Y= Ychromosome, X= X chromosome, y= yellow and yb=black, they thought Blue Boy was ybX/yY.
The fact that Blue Boy’s father was a yellow male made it probable that the y gene would be on the Y-chromosome rather than on the X. Mated to a tortoiseshell female, ybX/yX, a fertile ybX/yY tortoiseshell male could produce yellow males, tortoiseshell males and females, and black females as non-recombinant types ; and black males, tortoiseshell males and females, and yellow females as recombinant types. The production of all three genotypes in both sexes would be theoretically possible, which agreed with observation. About half the offspring from that type of mating would be tortoiseshells ; in fact, 14 out of 25, in fully sexed litters were tortoiseshell. The results of mating Blue Boy with yellow and with black females also agreed with their hypothesis of partial sex-linkage of yellow. Taken as a whole, the results suggested that linkage between the yellow locus and the sex-determining gene complex was fairly loose.
Various predictions could be made on the basis of their hypothesis ; for example, that those of Blue Boy’s sons which are yellow or black with regard to the y-locus should also be capable of producing tortoiseshell males.