GENETICS OF TAQPEP (TABBY PATTERN DEVELOPMENT)
MOLECULAR GENETICS OF SPOTTED CATS
In September 2012 researchers announced they had identified the gene that switches between mackerel and classic tabby patterns. This is the first "pattern gene" to be identified. In inheritance terms, mackerel pattern is the dominant "wild type" gene while classic pattern is due to a recessive gene. Cats with narrow stripes (mackerel tabby) have a working copy of the gene. The blotched pattern (classic tabby) occurs when a mutation turns that gene off. The researchers noted that cells in the black stripes (or spots in the case of cheetahs) know they are in a black stripe and remember that fact throughout the animal's life so that the pattern grows as the cat grows. The researchers looked at cells along the boundaries between light and dark stripes/spots. They analysed DNA samples from feral cats in northern California and the DNA of cheetahs, including samples from the blotched "king cheetah" (a recessively inherited "classic tabby" version of the cheetah). The research pointed to a gene they called Taqpep (Transmembrane aminopeptidase Q). Blotched house cats had mutations in this gene, while striped tabbies did not. The king cheetah also had a Taqpep mutation while the spotted cheetah had a normal version of the gene. The Taqpep gene produces an enzyme that diffuses outside of cells, interacting with other molecules. Another gene, called Edn3 (Endothelin3), led to the growth of dark fur rather than light fur and coordinated localized colour differences. Together, Taqpep and Edn3 produced the patterning: Taqpep establishes the type of pattern of stripes or spots in early development while Edn3 appeared to carry this on during growth.
Other genes, not yet identified, would determine further variations in the basic tabby patterns in domestic cats, for example broken stripes, round spots, marbled tabby (Bengal) or pale-centred blotches (Sokoke).
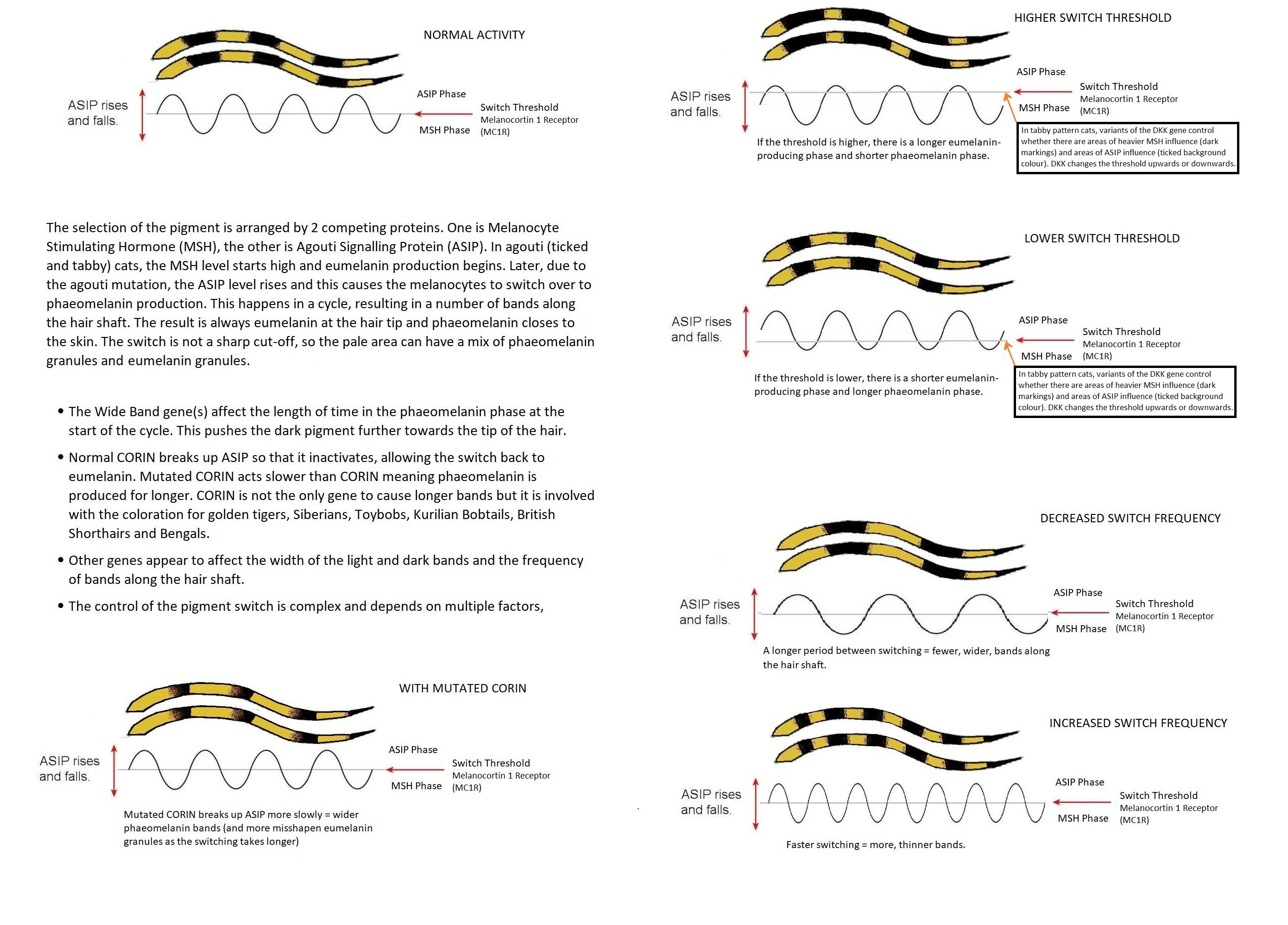
ACTION OF THE AGOUTI GENE - TICKED HAIRS
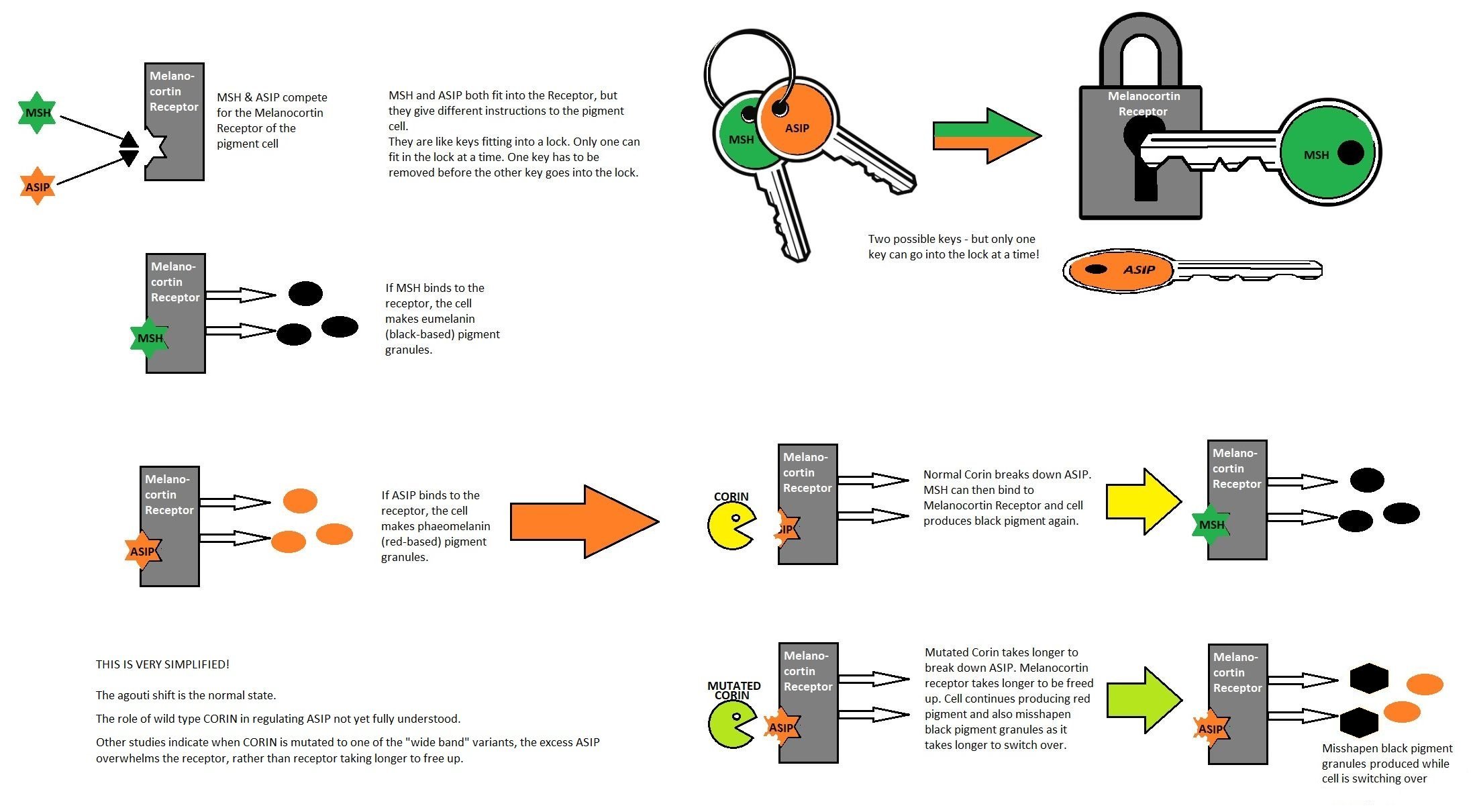
In 2011, the journal "Science" reported that the Agouti gene not only affects depth how deeply coloured the fur is, but is also involved in the formation of patterns. Researchers at Harvard University studied deer mice, which have dark backs and pale bellies. They showed that small changes in the activity of the Agouti gene during embryo development affects the distribution of pigment. Where the gene is highly active, it delays the maturation of the pigment-producing cells; this results in paler fur in adulthood, for example the pale belly found in many vertebrates. The researchers hope to find out how more complex patterns, such as tiger-striping and leopard-spotting, are formed and which genese are involved. In cats, the agouti gene determines the ticked (banded) pattern on the hair-shafts. Agouti areas of the body have ticked hair-shafts due to alternating bands of heavy and light pigment and are considered the "background colour". The solid patterns, such as tabby stripes, are non-agouti areas where the hair-shafts are not ticked.
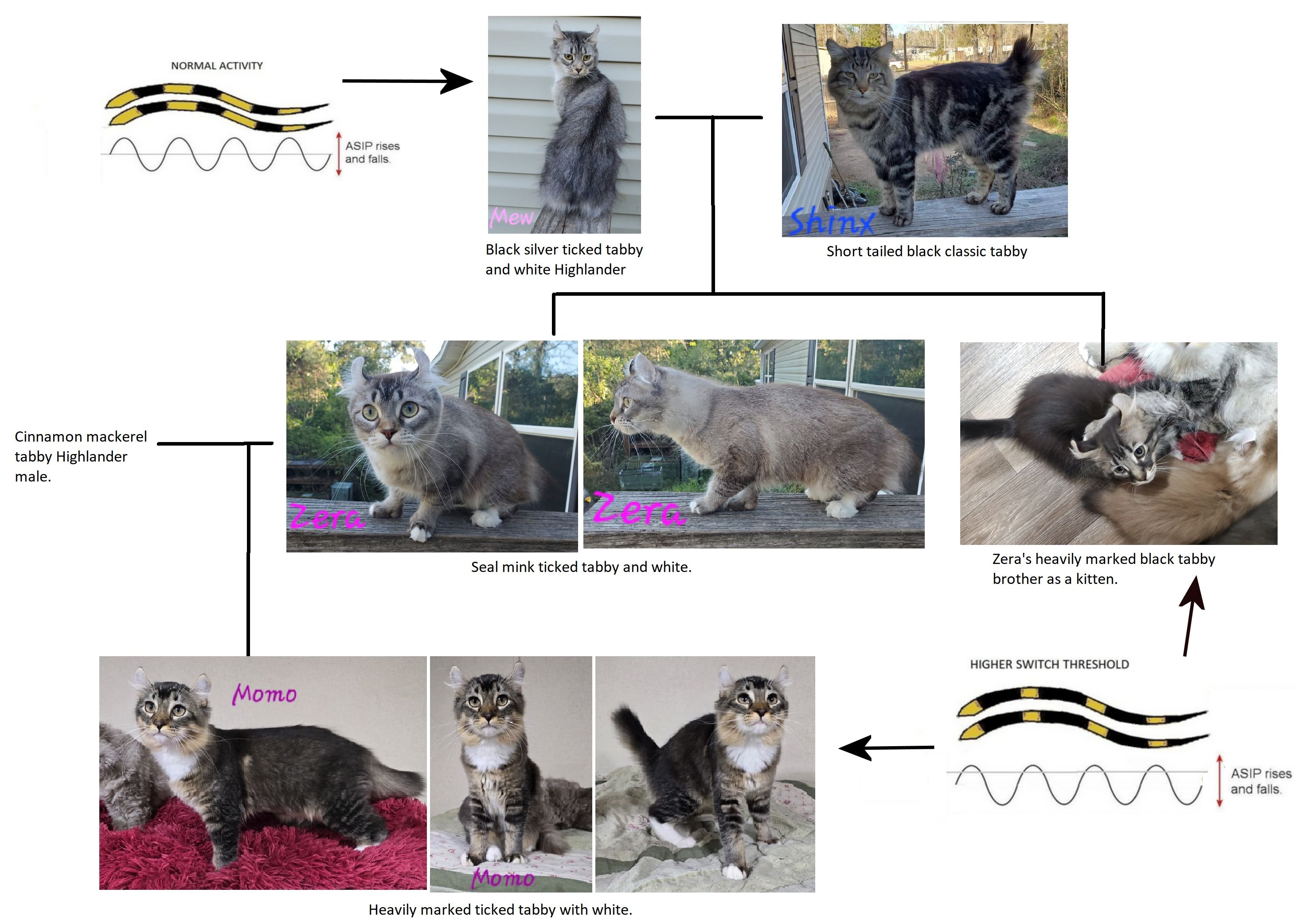
Krista Childers sent me examples of very dark tabbies in her Highlander lines following an outcross to a random-bred domestic. The first cats were Mew (black silver ticked tabby and white Highlander) and Shinx (short tailed black classic tabby). They produced Zera, a seal mink ticked tabby and white who was always darker than her mink sister. Her brother heavily marked mackerel tabby whose back appeared black as a kitten. Zera was bred to a cinnamon mackerel tabby Highlander and produced Momo, a heterozygous ticked tabby with homozygous classic "underneath" the ticked pattern. Luck of the draw has given Momo the genes for thick bands of black on the hair shafts and thinner bands of brown colour. The "switch" changing pigment deposition has been set "high" by the DKK gene. The "blacked out" pattern in her non-ticked tabby relatives is due to thick tabby markings and the darkening of the background colour. In ticked tabbies there is a "tide line" where the background colour shows up (this is seen more clearly in Abyssinians which have been selectively bred to maximise this trait). Combined with the heavy ticking and white markings it creates an interesting visual effect.
ACTION OF THE TAQPEP GENE (PATTERN DEVELOPMENT)
A summary of "DEVELOPMENTAL GENETICS OF COLOUR PATTERN ESTABLISHMENT IN CATS" by Christopher B. Kaelin, Kelly A. McGowan & Gregory S. Barsh in Nature Communications volume 12, Article number: 5127 (2021)
The researchers analysed cells from foetal skin of domestic cats to identify when, where, and how tabby patterns are established during foetal development. The foetuses were obtained from pregnant feral cats spayed during Trap-Neuter-Return programmes. Early in development, before the colour/pattern appeared, the researchers could identify stripe-like alterations in epidermal thickness related to expression of different genes. They found that the secreted Wnt inhibitor (a signalling molecule) encoded by 'Dickkopf 4' (gene, abbreviated to Dkk4) played a central role in this process, and is mutated in cats with the Ticked pattern type.
In mammalian skin and hair, melanocytes are uniformly distributed during development, and the amount and type of melanin produced are controlled later by paracrine signalling molecules within individual hair follicles. Paracrine signalling is a form of cellular communication in which a cell produces a signal to induce changes in nearby cells.
Step 1: Pattern element is set during foetal development.
Step 2: Pattern is implemented by paracrine signalling molecules produced within individual hair follicles e.g. producing light colour or dark colour hair
Step 3: Pattern element identity is maintained during hair cycling (growth-moult-regrowth) and cell division. This means individual spots or stripes present at birth enlarge proportionally as the infant grows.
In previous studies of domestic cats, the same researchers showed that Endothelin 3 is expressed at the base of hair follicles in tabby markings. More black-brown eumelanin is produced relative to reddish-yellow pheomelanin. This plays a key role in producing the tabby markings. Tabby markings are apparent in developing hair follicles before they produce hair i.e. colour patterns are established during or before hair follicle development.
Researchers divide embryonic/foetal into stages which they compare to the development stage in laboratory mouse. At stage 13 (analogous to mouse embryonic day 11), foetal skin is a uniform layer of epithelial (surface) cells, one cell thick, over a dermis that contains few cells. Approximately 16 days later at stage 16 (analogous to mouse embryonic day 15), before formation of hair follicles and other epidermal cell types, the epidermis is organized into alternating "thick" and "thin" regions that are different from the epidermal stratification (skin develops into layers) that happens later in development. By stage 22 (analogous to mouse post-natal day 4 6), there are well-developed hair follicles that can be categorized according to the type of melanin they produce. Dark markings contain mostly eumelanin, while light areas contain mostly phaeomelanin.
The researchers looked at natural genetic variation in Transmembrane aminopeptidase Q, (Taqpep) enzyme. Loss-of-function mutations in Taqpep cause the striped mackerel tabby pattern to expand into the blotched classic tabby. Both patterns are found in feral cat populations. Is the development of thick/thin epidermal areas at stage 16 influenced by Taqpep genotype. The thick epidermal regions of mackerel tabby embryos were organized into vertically oriented columns separated by larger thin epidermal regions. In classic tabby embryos, the thick epidermal regions in the flank and back of the neck are broadened. The thick/thin epidermis in the embryo resembled the tabby patterns of adults with corresponding genotypes (i.e. mackerel or classic) meaning the pattern is established before melanocytes enter the epidermis and before hair follicles develop. The alternating thick and thin regions arise from an earlier molecular pre-pattern.
The researchers looked at two Dkk4 variants called p.Ala18Val and p.Cys63Tyr that were only found in breeds where tabby markings were obscured: Abyssinian, Burmese and Siamese. These were strong candidates for "Ticked". All breed cats in which "Ticked" is required (Abyssinian, Singapura) carried the p.Ala18Val or p.Cys63Tyr Dkk4 variants, most being homozygotes, some being heterozygotes for the variants. In breeds requiring tabby markings (Egyptian Mau, Ocicat, Bengal), none carried these Dkk4 variants. In some breeds (Oriental Longhair, Oriental Shorthair) and in non-breed cats, which had either Ticked or non-Ticked phenotypes, these patterns correlated with the presence or absence of the p.Ala18Val Dkk4 variant.
A pedigree of Savannah cats found that the small-spot "servaline" phenotype was associated with the p.Ala18Val variant, but instead of suppressing the tabby markings as in the Abyssinian and Burmese, "Ticked" alters the number and size of tabby markings in Savannahs. A feral foetal skin sample heterozygous for p.Ala18Val variant showed a pattern very similar to the servaline pattern; the effect of "Ticked" was not to mask dark tabby markings (as in the Abyssinian and Burmese) but to affect pattern establishment so that regions that express Dkk4 in foetal skin, and the eventual adult markings, are smaller and more numerous.
In domestic cats, the servaline pattern is one of several examples demonstrating that the effect of Dkk4 depends on genetic background and genetic interactions. The different appearance of the Abyssinian and Burmese breeds and the interaction of the "Ticked" (Dkk4) and "Tabby" (Taqpep) genes support this. In the Abyssinian, "Ticked" is selected for its ability to draw attention to prominent hair banding patterns. However, in the Burmese breed, individual hairs are not banded due to a loss-of-function Agouti allele and Ticked has been selected for its ability to suppress tabby markings. Dkk4 is epistatic to Taqpep, such that the distinction between a Blotched and Mackerel phenotype can only be seen where "Ticked" is not present.
Selected References
Allen, W. L., Cuthill, I. C., Scott-Samuel, N. E. & Baddeley, R. Why the leopard got its spots: relating pattern development to ecology in felids. Proc. R. Soc. B: Biol. Sci. 278, 1373 1380 (2011).
Eizirik, E. et al. Defining and mapping mammalian coat pattern genes: multiple genomic regions implicated in domestic cat stripes and spots. Genetics 184, 267 275 (2010).
Eizirik, E. et al. Molecular genetics and evolution of melanism in the cat family. Curr. Biol. 13, 448 453 (2003).
Kaelin, C. B. et al. Specifying and sustaining pigmentation patterns in domestic and wild cats. Science 337, 1536 1541 (2012).
Kaelin, C. B. & Barsh, G. S. Genetics of pigmentation in dogs and cats. Annu. Rev. Anim. Biosci. 1, 125 156 (2013).
Kaelin, C. & Barsh, G. Tabby pattern genetics a whole new breed of cat. Pigment Cell Melanoma Res. 23, 514 516 (2010).
Lomax, D. & Robinson, R. Tabby pattern alleles of the domestic cat. J. Hered. 79, 21 23 (1988).
Lyons, L. A. et al. The Tabby cat locus maps to feline chromosome B1. Anim. Genet. 37, 383 386 (2006).
SOME OLDER THEORIES ON TABBY PATTERNS (BEFORE MOLECULAR GENETICS)
INHERITANCE GENETICS OF SPOTTED CATS
Early on when pattern inheritance was studied, it was believed that three alleles (gene variants) at the same locus (position on a chromosome) controlled the tabby pattern. Different pairings of these genes produced the following order of dominance in tabby patterns: ticked (Abyssinian) (Ta), Spotted (Ts), Mackerel (TM), and Blotched (Classic) (tb). However data from breeders indicated that mackerel and blotched were on one locus while ticked/non-ticked were at a different locus. Two copies of ticked completely masked the tabby pattern, one copy of ticked and one of non-ticked allowed stripes to show up on the extremeties while 2 copies of non-ticked allowed the tabby pattern to show through. Some other mechanism seemed to break up stripes into spots. More recent research (Eizirik, E, VA David et al) confirmed the 2 loci for ticked/non-ticked and mackerel/blotched and confirmed that a modifier at a 3rd locus can turn the mackerel pattern into well-defined spots. Three independent breeding lines in controlled environments were used:
Eizirik, E, VA David et al's paper proposed the symbol Ta with alleles TaM (mackerel, dominant) and tab (blotched, recessive) for the pattern-type locus; TiA (Abyssinian ticked) and Ti1 (non-ticked) for the other locus which can fully or partly mask the pattern. A name was not posted for the spotted modifier locus though Straede has previously posited Pmf" (breaks the pattern into spots) and recessive "pmu" (leaves the pattern unbroken).
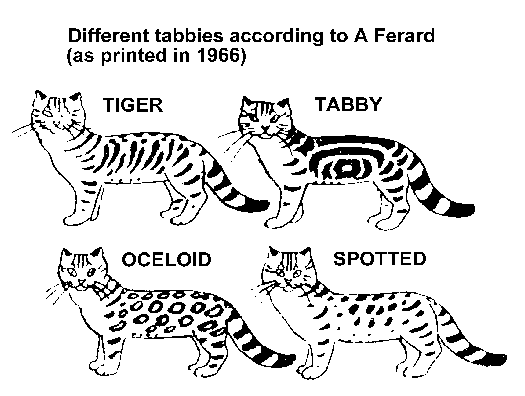
FERARD'S CLASSIFICATION OF TABBIES (HISTORICAL)
A Férard categorised tabbies as ranging from tabby (classic pattern), tiger, self (non-tabby), oceloid and spotted. Self indicates a solid colour. Agouti is not included. The oceloid pattern is intriguing as it suggests rosetting or clouding, a pattern only recently introduced into cats via hybridization with wild species.
Genes for creating or modifying spotted patterns have been introduced into the domestic cat from wild species. The Bengal breed (derived from hybrids with the Asian Leopard Cat) exhibits a more random type of spotting as well as the coveted rosette pattern (groups of spots) and a swirled marble pattern where domestic cat tabby and wild type spotting interact. The inclusion of the Bristol breed (a margay hybrid) into some Bengal lines may have added other genes for spots.
THEORY OF A PATTERN MODIFIER GENE
In 1992, Dr Truda Straede, originator of the Spotted Mist breed (now the Australian Mist, comprising Spotted and Marbled varieties) had concluded that there were only two tabby patterns - abyssinian and classic - and an independently inherited "pattern modifier" gene which she called "pm". The dominant "Pmf" form breaks the pattern into spots, the recessive "pmu" form leaves it intact (i.e. tabby). Pmf would interact with mackerel striping to give small spots or bars. It would interact with classic tabby to give larger, often oddly shaped, spots and bars. Since the Spotted Mist pattern size fell into large and small spots, she suggested that there was a second form of the classic gene which divided the large pattern concentrically into narrower bands. When mated together, "small pattern" cats never produced "large pattern" kittens. Since she had never observed very finely divided blotched patterns, either in purebreds on in random-bred cats, this suggested that the a gene for large pattern classic tabby (thl) was dominant to that for small pattern classic tabby (ths) and that the gene for the small form was only activated if the pattern modifier gene was active. Although Truda Straede had never seen any finely divided blotched patterns, such a pattern does exist. In the Sokoke, a natural Kenyan breed, the dark classic tabby markings do have a paler central area. The leading feline geneticist at the time, Roy Robinson, found Dr Straede's hypothesis interesting but disagreed with the overall conclusions regarding large pattern tabbies and small pattern tabbies.
There does appear to be a modifier which creates a spotted pattern out of an otherwise striped one. In classic blotched tabbies, the markings sometimes break up into larger round spots as found in the Ocicat and the Egyptian Mau. This suggests that spotted cats are not simply "broken tabbies" but have a separate gene for a spotted pattern or which interacts with the other genes to modify the pattern. This is especially the case where the spots are not vertically aligned as they would be in broken tabbies and where they are round rather than elongated.
OLDER & UNPROVEN THEORIES ABOUT TICKED CATS
There are a number of theories about how the ticked tabby pattern is modified. According to Albert C Jude in his book "Cat Genetics" (1955) ticking increases with age so that the adult tabby has more pronounced ticking than a kitten stage. In his investigation into the inheritance of tabby coat pattern, PW Whiting (1915) had tentatively considered ticking factors as a series of three alleles (3 alternative versions of the same gene) which he called "much ticking" "little ticking" and "non-ticked". Whiting also considered 3 alleles of the banding factors (number of bands on each hair), calling them "lined" "striped" and "blotched". Both sets of alleles above are named in the order of dominance suggested by Whiting.
While researching the relationship between classic, mackerel and ticked tabbies and their relationship to silver shaded cats, American Shorthair breeder Carol W Johnson suggested the existence of a "Chaos" gene which disrupts the normal striped pattern. In ticked tabbies, Chaos (along with Johnson's theorised "Confusion" gene and Cathy Galfro's suggested "Erase" gene) might eradicate residual markings. In a classic tabby, Chaos causes an intermixing of the light striped areas into the black and vice-versa. The banding of each hair is normal, but the hairs themselves occur in the wrong places. The effect she describes is more clearly seen in the Sokoke, which has a modified tabby pattern. An alternative mechanism for this was theorised by Australian Mist breeder Truda Straede with what she referred to as the "finely divided tabby pattern". Currently, genes such as Chaos, Confusion and Erase are theoretical. It is plausible that some of the many polygenes affecting hair ticking and banding do indeed have the effects attributed to these genes.
Johnson also noticed that unpatterned tabbies, closely resembling ticked tabbies, sometimes occur in purebred where the conventional ticked tabby pattern is not present and where outcrosses to ticked (Abyssinian type) cats are not acceptable. They occurred when a Shaded cat was crossed with either a Classic tabby or with a solid/self carrying classic tabby. She described this phenomenon and formed a hypothesis based on observations and test matings. At the time, the Abyssinian-type ticked tabby was believed to be an alternative to mackerel tabby or classic tabby i.e. there were three versions of the same tabby gene. What she described was that mackerel and classic tabby were two versions of the same gene while "Unpatterned" tabby (i.e. Ticked Tabby or Abyssinian-type) was an entirely different gene carried elsewhere on the chromosome. It masked out any other tabby pattern present in the cat's genes. In addition, a cat with two copies of the Unpatterned (tabby gene (ticked tabby gene) had fewer residual markings (barring on legs, chest and head) than a cat with only one copy of the gene. A cat with no copies of the Unpatterned gene allowed any other tabby pattern present to be expressed. She also suggested other genes which remove residual markings, noting that some ticked tabbies show "ghost" striping which superficially resembles a mackerel tabby, but is genetically different, perhaps being caused by presence or absence of an as yet unidentified gene.
Johnson considered that the this Unpatterned Tabby gene was present in Shaded Silver American Shorthairs as mating Shaded Silver to Classic Tabby sometime produced ticked tabbies, which were not recognised by the breed society. It is possible that it did enter the Shaded colours at some point in history, but it is masked by the Shaded pattern and hence only manifests in kittens born from matings with non-Shaded cats. According to Johnson, the occurrence of unpatterned Chinchilla kittens suggests that the ticked tabby gene may be present in Persians although I have seen no data regarding ticked tabby offspring from matings of Chinchilla/Shaded Silver to Classic Tabby.
|
|
Non-agouti (Self) |
Agouti Mackerel/Classic |
Heterozygous Agouti Unpatterned (Ticked) |
Homozygous Agouti Unpatterned (Ticked) |
|
Inhibitor |
Smoke |
Silver Tabby |
Shaded/Chinchilla |
Shaded/Chinchilla (with no breakthrough pattern) |
|
No Inhibitor |
Self |
Patterned Tabby |
Ticked (Abyssinian) Tabby |
Ticked (Abyssinian) Tabby |