BASIC GENETICS AND CELL DIVISION
This is a very simple guide to inheritance provided as an aid to understanding other Messybeast genetics articles. If you are interested in more scientific details or understanding inheritance in greater depth, a search engine will find plenty of websites that explain genetics in more detail. An overview of cell division is provided as background into how each parent contributes half towards the offspring's genetic complement, how genes are shuffled and how mutation can occur during cell division.
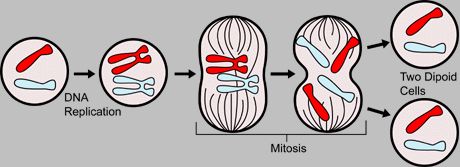
A cat s body cells all have 2 sets of chromosomes (called diploid), one set inherited from each parent. When body tissues are renewed, the cells undergo cell division to produce genetically identical cells with the same number of chromosomes as the parent cells. This is called mitosis.
When germ cells are produced, the ova (egg cells) and sperm cells must only have one set of chromosomes (called haploid) in order to make a diploid embryo when an egg is fertilised. This process is called meiosis and is part of sexual reproduction in cats (and in humans, dogs etc). This is called meiosis and allows genetic traits to be shuffled so that the offspring have a different set of traits to either parent.
The following description of cell division is greatly simplified and provided as an aid to understanding how genetic traits are shuffled and how mutation occurs. For more detailed information on the different stages and processes involved in each type of cell division, there are many resources on the web.
MITOSIS SOMATIC (BODY) CELL DIVISION
In mitosis, one cell diploid divides into 2 daughter cells while preserving the diploid chromosome count. Each daughter cell is genetically identical to the parent cell and to the other daughter cell (barring mutation due to random transcription errors).
A diploid cell has a full set of chromosome pairs. Each pair contains one chromosome from each parent. These chromosome pairs are not identical, but are homologous (equivalent) chromosomes as they contain different alleles of genes. For example, one particular locus (location a bit like a house number on a street) on one of the father's chromosomes may pass on black fur. The same locus on the mother's chromosome may pass on grey fur.
Firstly, the parent cell must make copies of each chromosome. The two identical copies are called sister chromatids and are attached together at a specialized region of the chromosome called the centromere. At this point, the cell contains twice the normal amount of genetic material. The chromosomes then align themselves in a line across the cell. The chromotids are pulled apart and one chromatid, now called a chromosome again, migrates to each pole of the cell and the cell itself splits into 2 cells with each cell having the normal amount of genetic material.
When DNA is replicated, each chromosome makes an identical copy of itself. One copy will end up in each daughter cell so that the daughter cells have the same combination of genetic material that the parent cell had. Homologous chromosomes are kept together. In the diagram, the parent cell has one red and one blue chromosome. Both daughter cells will also get one red and one blue chromosomes.
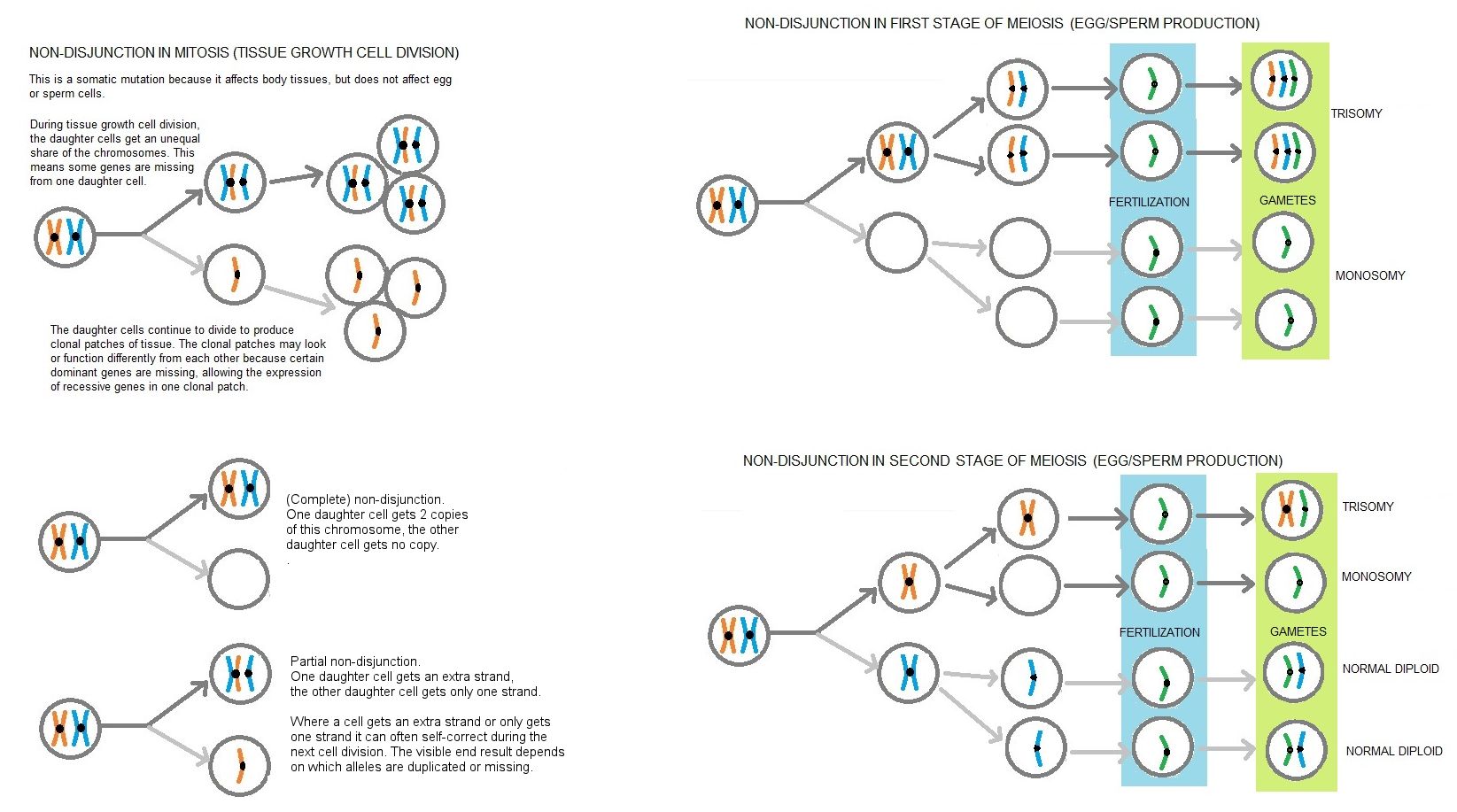
Sometimes there are errors in mitosis. This affects not only the immediate daughter cells, but the future offspring of those daughter cells. For example, in non-disjunction, the chromatids fail to separate and one daughter cell gets both sister chromosomes while the other daughter cell gets none. This can result in cancer. Chromosomes may also become damaged with parts broken off, wrongly copied, duplicated or reattached wrongly (looped or reversed). The effects range from no noticeable effect to lethal (especially in the embryo which is rapidly dividing and where early mistakes are multiplied many, many times). These mistakes are called somatic mutation because they affect the body cells and not the germ cells.
MEIOSIS PRODUCTION OF GERM CELLS (OVA AND SPERM)
Meiosis produces four genetically varied gametes (egg cells or sperm cells) out of a diploid cell. A diploid cell has a full set of chromosome pairs. Each pair contains one chromosome from each parent. These chromosome pairs are not identical, but are homologous (equivalent) chromosomes as they contain different alleles of genes. For example, one particular locus (location a bit like a house number on a street) on one of the father's chromosomes may pass on black fur. The same locus on the mother's chromosome may pass on grey fur.
First, the chromosomes are replicated. Each chromosome comprises two identical chromatids joined together by a stretch of DNA called the centromere. Meiosis I, the first round of division, separates homologous chromosomes. Meiosis II, the second round of division, separates sister chromatids. The end result is 4 haploid cells each containing slightly different mixes of genes.
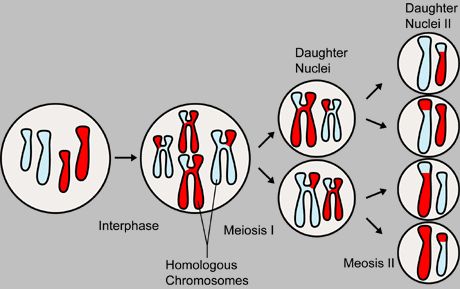
During meiosis I, the chromosomes are replicated and genes may be shuffled from one chromatid to another. The chromosomes pair up together for a short time during which chromosomal crossover occurs a gene on one chromosome may get swapped for the gene on the corresponding chromosome (a bit like 2 people crossing the road in opposite directions at a pedestrian crossing). In the diagram, there are 2 chromosome pairs (big chromosome and little chromosome). The red part was inherited from the father and the blue part from the mother, so the pairs of chromosomes in the offspring have one red and one blue chromosome. Crossing over of genes is shown as red and blue patches. This results in 2 diploid cells, each containing a slightly different mix to the original parent cell.
During meiosis II, each of the 2 diploid cells splits into 2 haploid cells. It skips the stage of replicating chromosomes and goes straight into the division stage. As a result of this, the cells have half the number of chromosomes. These are the ova (egg cells) and sperm cells, known collectively as germ cells. When they meet up in a fertilised egg, the resulting embryo will have the normal diploid number of chromosomes.
If the number of chromosomes was not halved, each generation would have twice as many chromosomes as its parents. Some hybrids are triploid (3 sets of chromosomes) or tetraploid (four sets of chromosomes) but are usually sterile because meiosis cannot occur. Having three or more sets of chromosomes is known as polyploidy and can result in developmental abnormalities. Sometimes there are errors during DNA replication parts of the DNA may be lost, damaged, duplicated or reversed. This is mutation. If a germ cell with mutant DNA fertilises (or is fertilised by) another germ cell and results in an embryo, the embryo inherits the mutation. This can range from not noticeable (recessive genes or "redundant" DNA) to cosmetic (fur colour) to lethal (body structures are not properly formed). Mutations which are not noticeable in the offspring may show up in later generations if two identical mutant genes meet up.
Sometimes chromosomes do not separate properly during meiosis I or chromatids do not separate properly during meiosis II. As a result the germ cells have either more or less genetic material than normal. A chromosome might be duplicated or absent. The most obvious instances in cats are when one of the sex chromosomes is duplicated or missing. Normal females are XX and normal males are XY. An XXY cat is a sterile male with a tendency to giantism or overweight; this is one cause of the uncommon tortoiseshell male cat. An XO (Turner syndrome) cat is a sterile undersized female, often with heart problems.
CHROMOSOMES, GENES AND ALLELES
A chromosome is a length of DNA. In cats, as in humans, autosomes (chromosomes other than X or Y) occur in pairs. Cats have 19 pairs of chromosomes. The matching strands of DNA form a double helix. The exception to this are the sex-determining chromosomes X and Y. A gene is a stretch of DNA that, in simple terms, encodes for a trait such as long or short fur. On the autosomes, genes occur in pairs - one on each strand. These genes may be identical or they may be slightly different variations of the same basic gene. The variants are called properly called alleles of genes, but for simplicity I shall call them genes (they should also be called "gene for black", "gene for grey" etc, but I shall refer to them as "black gene", "grey gene").
The "locus" of a gene refers to the place it occupies on the chromosome. Think of the chromosome as being like a street, the gene being a house and the locus being the house number. The street has 2 sides and the houses opposite each other may be identical or different.
Most cells have pairs of chromosomes (called diploid). The exceptions are egg cells and sperm cells which have a single set of chromosomes (called haploid). This is so that egg and sperm cells can fuse into a fertilised egg that has the correct number of paired chromosomes, one set from the mother and one set from the father. The offspring gets half of its genes from each parent and will be genetically different from each. Working out which animals to mate together to achieve the desired results in the offspring is both a science and an art (and the subject of many full-length books).
SIMPLE INHERITANCE (MENDELIAN INHERITANCE, CLASSICAL INHERITANCE)
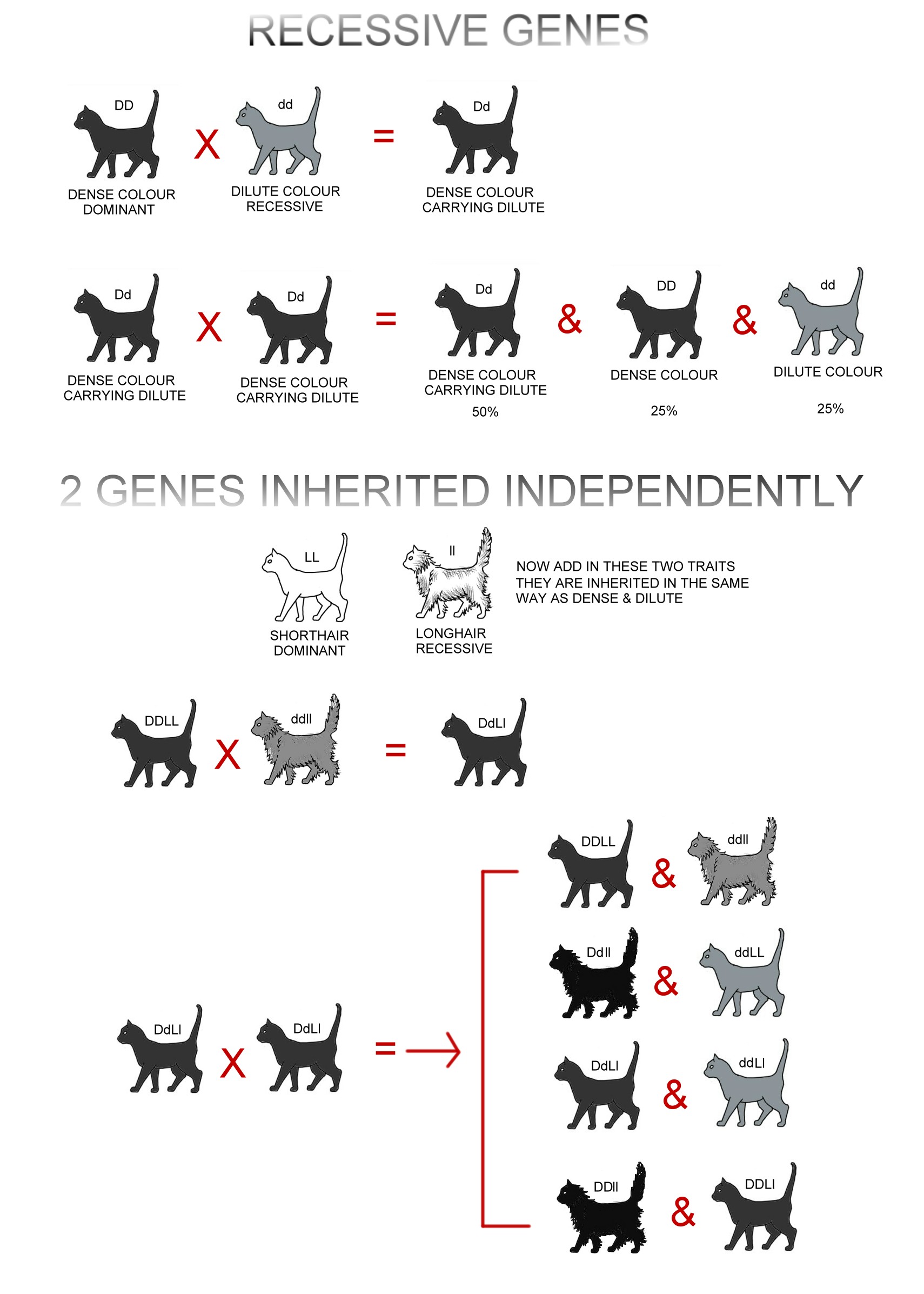
Named after the studies of Gregor Mendel, simple inheritance relates to a trait governed by a single gene. In cats, the inheritance of black and blue (grey) is an example. Genes on the autosomes (the chromosomes other than the X and Y sex chromosomes) are inherited in pairs, one copy from each parent. If a cat inherits 2 blue genes, it will be grey. If it inherits 2 black genes it will be black. If it inherits 1 black gene and 1 blue gene it will be black but can pass on the blue gene to its offspring.
The black gene is dominant - this gene will always be expressed (show up in the phenotype i.e. outward appearance) even if only 1 copy is present. The blue gene is recessive and will only be expressed if 2 copies are present. An animal where both copies of the gene are the same is homozygous for that trait. An animals where the copies are different is heterozygous for that trait.
The blue gene is recessive. It is only expressed if the cat didn't inherit a black gene. Blue cats are homozygous for the blue colour. Its genotype (genetic make-up) is the same as its phenotype unlike a black cat whose genotype can be either heterozygous or homozygous.
By convention, a dominant gene is shown in uppercase and a recessive gene is shown as the same letter, but in lowercase. I have also shown the letters in colour.
|
|
Black homozygous cat (BB) mated to blue homozygous cat ( bb) |
|
|
Black heterozygous cat (B b) mated to black heterozygous cat (Bb) |
||
|
|
B |
B |
|
B |
b |
|
|
b |
B b |
B b |
B |
BB |
B b |
|
|
b |
B b |
B b |
b |
B b |
bb |
|
In some cases, neither gene is dominant. The genotype Xx would then give a phenotype (appearance) intermediate between XX and xx in the same way that pink is intermediate between red and white. In the 19th Century, breeders believed all genes worked this way and called it "blending".
DOMINANT GENES, RECESSIVE GENES, EPISTATIC GENES
A dominant gene is always expressed even if only one copy is inherited. A recessive gene is only expressed if two copies are inherited.
Another type of gene sometimes called "dominant" is an "epistatic" gene. This is a gene that resides at a different locus of the chromosome or on a completely different chromosome and it masks the effect of certain other genes. An example in cats is "dominant white" (properly called "epistatic white"). When present, dominant white masks out any other colour genes. The cat has a white phenotype but also has the genes for other colours and passes on those colour genes to its offspring.
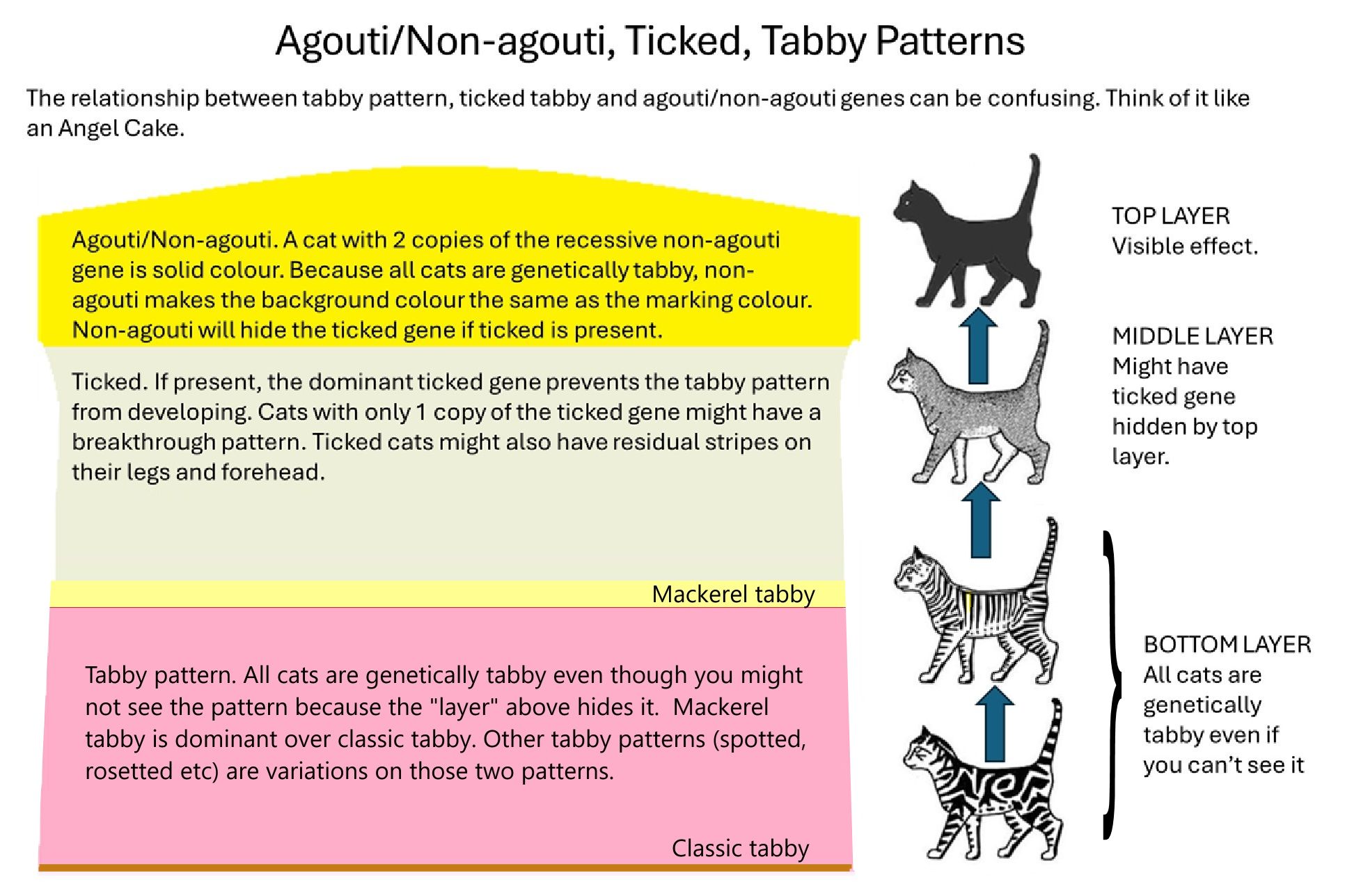
Orange (or red) is another epistatic colour as it influences the non-agouti gene. Non-agouti (solid colour) cats that also have the orange gene will turn out tabby. Even the so-called solid red (red self) cat has tabby markings, but breeders have worked to decrease the contrast between background and marking colours to give the impression of no markings.
Agouti (Abyssinian tabby) is epistatic over other tabby patterns. Until recently it was thought to be an allele of tabby. Recently it was found to be on a different locus.
Although genes are inherited in pairs, there may be more than 2 variants of a gene carried by different individual. Some of those genes are dominant, some are recessive and some are co-dominant (neither dominant nor recessive). An example in cats is the colourpoint series. This is a form of temperature dependent albinism where the cooler parts of the cat (legs, tail, ears, muzzle) are darker than the warmer parts (torso). The colourpoint gene is recessive to the solid colour gene.
|
Dominant |
Recessive(s) in ascending order of dominance |
Note |
|
C - solid colour |
c - pink-eyed albino These are co-dominant with each other:- |
cs/cb = Tonkinese pattern (mink) |
There are many possible combinations because some genes are more recessive than others!
* If C (full colour) is inherited, the cat will be solid colour and any other genes will be hidden.
* If C is not inherited, the other combinations will show up as below:
* If cs is inherited, the cat is colour point and any c or ca genes are hidden
* If cb is inherited, the cat is sepia and any c or ca genes are hidden.
* If neither cs nor cb are present, ca is inherited, the cat is blue-eyed albino (recessive white) and the c gene (if present) is hidden.
* If none of cs, cb or ca are present, 2 c genes are inherited, the cat will be pink-eyed albino.
There is one complication: cs, cm and cb are co-dominant (dominant over other genes, but not dominant over each other). If a cat inherits one cs + one cb, the pattern is intermediate between colourpoint and sepia and is called mink. The Tonkinese breed was created by breeding Siamese and Burmese cats together. In the same way cs + cm creates intermediate form Siamocha, while cb + cm creates intermediate form Burmocha.
Inbreeding (breeding together closely related individuals) tends to bring out recessive genes because the animals carry similar combinations of genes and there is more likelihood of any hidden genes meeting up and, therefore, showing up. Inbreeding does not create recessive traits, it just increases the probability of recessive genes being manifested.
A gene series must occupy the same loci on pairs of chromosomes. Some websites based on older texts incorrectly show tabby as a series of genes in the following order of (most dominant first): Abyssinian (ticked) Tabby, Mackerel Tabby, Classic Tabby. This is incorrect because Abyssinian Tabby is on a different locus.
Most traits are inherited separately from each other because the genes occupy different loci (plural of locus) on the chromosomes. Some traits are inherited together because the genes are located close to each other and tend to be passed along together. Think of it as being like terraced houses on a street - you can't move one of the terraced units without moving all of them because they share some walls.
Two grey cats mated together produce more grey cats because they only have the recessive genes. Two black cats mated together might produce grey kittens if both parents carry a hidden recessive grey gene. Looking at the pedigree of the parents, it will turn out that each cat has a grey ancestor. The hidden recessive gene has been passed down over several generations, but only shows up when a kitten inherits 2 recessive genes and no dominant gene to mask it. This is sometimes called a throwback because the kitten is a throwback to an earlier ancestor.
Some genes are carried on the X and Y sex chromosomes. Because these chromosomes do not form pairs in the same way as the autosomes, the traits affected show up unequally in males (XY) and females (XX). These traits are known as sex-linked. In cats, the tortoiseshell pattern is sex-linked (this includes the calico pattern which is simply tortoiseshell with the addition of the white spotting gene) and the majority of tortie cats are female. Male torties are uncommon and are due to developmental or genetic abnormalities.
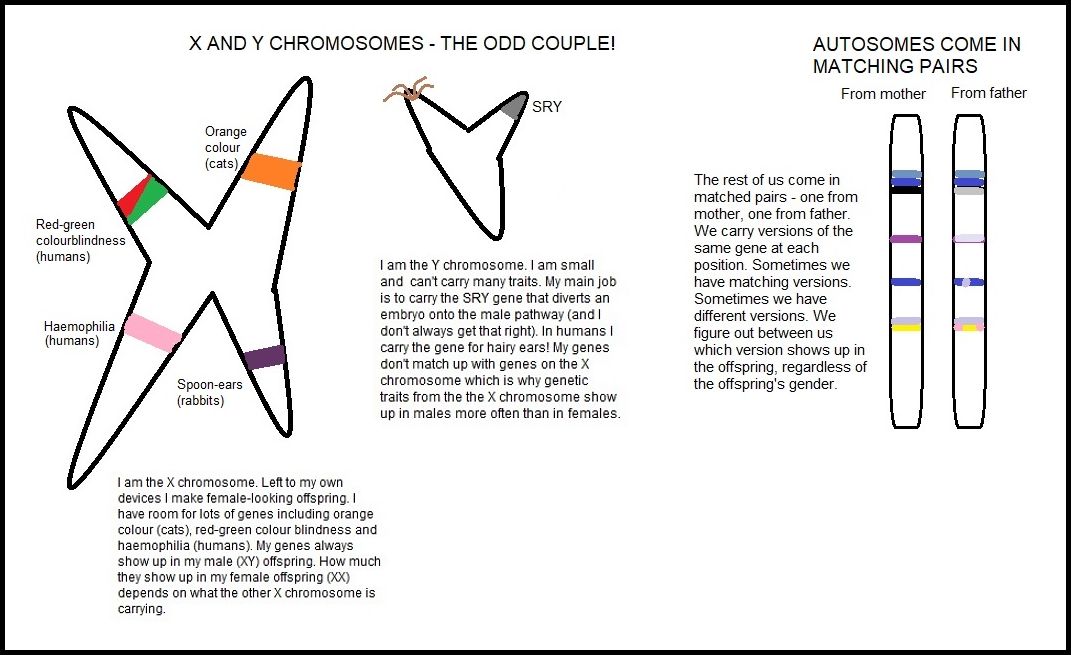
The orange gene (O) changes black pigment into a reddish pigment. The orange gene is carried on the X chromosome. A normal male cat has XY genetic makeup so he only needs to inherit one orange gene for him to be a ginger cat. A normal female is XX genetic makeup. She must inherit 2 orange genes to be a ginger cat. If a female inherits only one orange gene, she will be tortoiseshell and will be heterozygous for both orange and black. (The actual formation of orange and black patches in a tortie female is because, early in embryo development, some cells randomly activate the orange gene from one X chromosome while others activate the black gene from the other X chromosome. This happens in all cells, but only shows up visibly in skin cells as these produce pigment.)
More accurately speaking, O = "epistatic orange" and o = "non-orange" meaning that non-orange allows whatever colour is carried on the other chromosomes to be displayed (black, blue, chocolate etc). For simplicity, I am only considering red/black tortie and have referred to o as "black". "Orange" is also known as "red" depending on the age of the text you are reading. A "-" means there is no corresponding gene because the sex chromosomes do not form a pair.
|
|
GENES FROM TORTIE FEMALE |
|
|
GENES FROM TORTIE FEMALE |
||
|
GENES FROM RED MALE |
O (orange) |
o (black) |
|
GENES FROM BLACK MALE |
O (orange) |
o (black) |
|
O (red) |
OO Red female |
Oo Tortie female |
|
o (non-red [black]) |
Oo Tortie female |
Oo Black female |
|
No gene (Y chromosome) |
O- Red male |
o- Black male |
|
No gene (Y chromosome) |
O- Red male |
o- Black male |
![]()
Some traits are influenced by multiple genes carried on different loci of different chromosomes. Ginger cats vary from show-quality deep red through to sandy yellow depending on which polygenes are modifying the basic colour. Some genes intensify the colour while others weaken it. All ginger cats have the O (orange) gene, but the different mix of polygenes causes the great variation in shade. Likewise, grey cats range from a dark grey through to much paler versions.
Mating the palest ginger cats together means some offspring may inherit several genes that lighten the overall colour. In exhibition quality red cats, breeders have mated together the deepest coloured cats over many generations to produce richly coloured cats. These cats have the same gene for orange that is found in sandy coloured non-pedigree cats, but they also have more of the polygenes that intensify the colour. Ginger cats vary from deep red through to sandy yellow depending on what polygenes are modifying the basic colour.
CO-DOMINANCE AND INCOMPLETE DOMINANCE
Co-dominance, where two genes are equally dominant and the result of having one of each is a halfway point, has already been mentioned. Both genes are expressed (at least partially). The Tonkinese mink colour is intermediate between the Burmese sepia and Siamese colourpoint colour and pattern because the cb and cs genes are neither dominant to, nor recessive to, each other.
In incomplete dominance (partial dominance), a heterozygous individual (one who has both the dominant gene and the recessive gene) also has an appearance intermediate between a homozygous dominant individual and a homozygous recessive individual. The result is a blend of traits. If two individuals with an incompletely dominant trait are bred together, there will be a mix of offspring. Some will have the dominant trait, some will have the recessive and some will have a blend of traits.
|
R = dominant |
R |
r |
|
R |
RR |
Rr |
|
r |
Rr |
rr |
Penetrance means the extent to which a gene is expressed. Even a dominant gene may not be fully expressed because of the effects of other genes and of the environment. A highly penetrant gene is almost always expressed regardless of other genes or the environment. A low penetrant gene is only sometimes expressed. Highly penetrant genes are easily noticed and the mode of inheritance (dominant or recessive) more easily worked out. Some genes have age-related penetrance and start showing up as the individual ages.
Classic polydactyly in cats is dominant with incomplete penetrance. Cats with the normal number of toes sometimes produce polydactyl offspring. Barring random mutation, the parents are genetically polydactyl, but other genes or environmental factors affecting embryo development meant they didn't develop the extra toes. In this case, although the trait was suppressed in the parents, it showed up in the offspring. Cases of incomplete can resemble a recessive trait until enough breeding statistics are gathered.
The process of duplicating DNA in a cell can result in errors. These are comparable to spelling errors when copying out a chunk of text. Mutation is where a change to the DNA affects a gene and the change shows up in individuals inheriting that change. Over many generations, mutations accumulate. Mutations in body cells are called somatic mutations. They may show up in the individual in whose body they occur, but they are not passed on to the next generation (for example some skin cancers). Mutations in sperm or egg cells are germ-line mutations. They are passed onto the offspring.
Mutations can affect physical traits such as colour, fur type etc. Some cause devastating damage and the individual dies or is deformed. Others have no apparent effect. Some have a cosmetic effect such as rex (curled) fur. Some have a beneficial effect, better suiting the individual to survival in a changing environment.
During cell division, each daughter cell should get one copy of each chromosome. Sometimes this goes wrong and one daughter cell gets two copies, or one-and-a-half copies , while the other daughter cell gets no copy, or half-a-copy. The effect of this depends on what is on the chromosome and how it is shared out. The cells with too much or too little might simply die. Sometimes the cell can compensate during the next cell division. Or they may give rise to tissues that work normally, but are genetically different from other tissues. One example can be be found in black cats carrying one dominant allele for dense (black) colour and one recessive allele for dilute (blue) colour. If cell division does not happen properly, both copies of the dominant allele can end up in one daughter cell and one or both recessive alleles (but no dominant black allele) can end up in the other daughter cell. If both daughter cells continue to multiply, then the animal will have a mostly black coat with a fairly large blue area.
Genes made up of nuclear DNA are not the only form of inheritance. In animals, "throwbacks" are usually attributed to carried recessive genes. However, RNA may be responsible in some cases. "Paramutation" is the phenomenon where the characteristic governed by a gene is "remembered" and shows up in later generations, even though the gene itself is no longer present. Some primitive organisms, such as viruses, store their genetic material in RNA rather than DNA. In higher organisms, RNA still plays an important part in protein synthesis in the body so it is not surprising that RNA can influence how traits are passed on, expressed or suppressed.
Recessive genes are not the only way animals can pass on unexpected traits to their offspring. Mice could inherit a trait even if they didn't inherit the genes for the trait. In experiments for spotty-tail mice, the normal Mendelian rules are:
2 normal Kit genes = non-spotty tail
1 normal Kit gene + 1 mutant Kit gene = spotty-tail
2 mutant Kit genes = offspring that die shortly after birth
The mutant form of the Kit gene is dominant. Mice with non-spotted tails therefore have 2 copies of the normal Kit gene and, when mated to other non-spotty tail mice, should not produce offspring with spotted tails.
Researchers at the French Institute of Health and Medical Research and the University of Nice-Sophia Antipolis found that some mice born with two normal Kit genes had a spotted tail. They suggest that RNA (a polynucleotide related to DNA) passed on the trait to the offspring. RNA plays an important role in protein synthesis and other chemical activities in the body. The researchers found that the mutant Kit gene produced large amounts of messenger RNA which accumulated in the sperm of the mice and passed into the fertilised egg. Once in the fertilised egg, the normal Kit (non-spotted tail) gene is silenced and the embryo develops a spotted tail. RNA from the sperm of a mutant mouse gave the same result when injected into a fertilised mouse egg.
WHY GENETIC ENGINEERING ISN T SO SIMPLE
A gene does not control one trait only. A gene controls how a protein is made or it gives an instruction in what to do next . Proteins are the building blocks of our bodies. Genes are a bit like a recipe or an instruction list.
For example we are making house-bricks. The usual recipe makes strong pink house-bricks. We want to make some that are extra-red so we change the recipe (= mutation) to have more red dye. But the extra-red bricks are only half as strong so the house is a nice red but it is not strong. Or maybe we want to make blue bricks. We change the original recipe (= mutation) to use blue clay instead of pink clay. But the side-effect of changing the recipe is that the bricks can t be baked in at the usual temperature in the kiln (oven) because they don t go hard at that temperature. These bricks are useless for building houses.
The original recipe makes great bricks, but the recipes to give extra-red bricks or blue bricks change other things, not just colour. So you change an ingredient in the recipe to make strong bricks that bake at the regular temperature in the kiln. But this means the bricks will always come out black no matter what dye or clay you use! After lots of trial and error, you realise that only the original pink house-bricks work out okay. Any other recipe gives problems (so you want different colours you will have to use paint after the house is built).
Genes are like this - when you change or add a gene, it affects more than one thing because it changes a protein (which acts as a building brick or an instruction) that is used all over the body. Or you can think of it like changing the lock on your house door so that the key doesn t fit the new lock and you can t go any further into the house you get stuck at the instruction put key in lock and you can t get any further . Or you could think of it like a book where you insert a chapter from a completely different book: the story doesn t make sense!
In cats, the gene mutation that gives folded ears affects the ear cartilage. Genes need to be in pairs, and if one copy is the fold-mutation gene the cat has folded ears, but it also has side effects that can affect the bones and joints in the hind legs and tail make walking painful.
Maybe you add a jellyfish gene that makes a cat glow bright blue in the dark. The gene changes a protein that affects pigment. But that protein might also be used as an instruction in eye development in the embryo. The embryo can t develop any eyes because it the protein is wrong and the next instructions can t be followed. The blue-glow-in-the-dark gene works okay in the jellyfish s instruction set, but not in the cat instruction set. This is why can t just add or change a gene and get the effect you want you also get side effects because the genes change instructions on how to build the body, or they change proteins so that instructions get messed up.
Epigenetics ("in addition to genetics") refers to changes in gene expression during cell division or between generations where there aren't any mutations in the actual genes. The changes in the individual are due to the way the genes are expressed or silenced. A crude analogy would be the role of punctuation in a sentence: the words in the sentence are the same, but the punctuation gives them different meanings e.g. "Woman, without her, man is useless" and "Woman without her man is useless". Epigenetics involves modifications to DNA and associated proteins (analagous to the punctuation marks). It is an important part of cell differentiation in embryos where some genes are activated and some suppressed and these changes remain stable when each type of cell multiplies (a cell that becomes a heart muscle cell will divide to form more heart muscle cells). Several types of epigenetic inheritance systems may play a role in cell memory i.e. the cell remembers to make the right type of daughter cells. Some epigenetic features show transgenerational inheritance i.e. are inherited by the subsequent generation(s).
Epigenetic processes include paramutation, imprinting, gene silencing, X-chromosome inactivation, maternal effects, cancers and developmental abnormalities. Epigenetics imposes technical limitations on cloning due to the timing of turning genes on and off. This is a very simplified overview of a very complex topic.
Epigenome - the overall epigenetic state of a cell.
epigenetic code - the set of epigenetic features that create
different phenotypes in different cells.
DNA methylation and chromatin remodelling
Genes are carried on chromosomes. Chromosomes are made of chromatin
which is made of DNA and histone proteins. The outward appearance of an
individual is affected by which of its genes are transcribed (read and
acted upon). Genes can be transcribed in different ways and heritable
transcription states can result in epigenetic effects. Unlike gene
mutation, these effects are reversible and may be reset in the next
generation. Gene expression is regulated in several ways, for examply
by chromatin remodelling. Chromatin remodelling begins with
modification within the histones or by adding methyl groups to specific
areas of the DNA. These changes can act as templates or can
propagate.
The tails of the histones can be modified in different ways including acetylation, methylation and ubiquitylation. These changes affect how the DNA folds itself up or opens itself out so that enzymes can transcribe the genes it carries. Different changes can attract or repel the enzymes that do the transcription so some genes are expressed and others aren't. It's a bit like having a book where some pages are sealed together to prevent reading and where the book falls open at certain pages to catch the reader's eye.
Methylation may help silence "junk DNA" (redundant or repeated genes) and in some circumstances might result in permanent genetic changes (gene mutations). DNA methylation patterns are influenced by the environment and can be established and modified by environmental factors. It is involved in proper development of the embryo, imprinting and, in females, X-inactivation (X-inactivation gives rise to tortoiseshell cats).
Chromatin remodelling means some regions of a chromosome can take on stable and heritable alternative states without any change to the DNA. This means the same gene could be expressed in two different ways.
DNA methylation and chromatin remodeling is important in cell differentiation during the development of multicellular animals. While the actual genome remains the same, the cells differentiate into many different types and create different tissues and organs in the body. Morphogens activate or silence genes in an epigenetically heritable manner. This epigenetic inheritance means differentiated cells have a memory and when they divide they make more of the same type of cell e.g. a liver cells makes more liver cells (stem cells are an exception).
RNA effects
Genes are turned on in response to a stimulus. Some genes, after
being turned on for the first time, transcribe products that keep the
genes turned on. This may involve producing different forms of RNA.
When the cell divides, its descendents inherit the turned-on gene (even
though the original stimulus is absent) and the different forms of RNA.
When eggs are formed within the mother, it inherits large quantities of
RNA from her. This affects how the genes are switched on if the egg
develops into an embryo (maternal effect phenotype). Sperm carries
less RNA, but this can also have an epigenetic effect on the
offspring.
So, it is not just a question of which genes the offspring inherits, it is also a question of which genes are turned on and whether a gene can be transcribed in more than one way and whether a form of RNA associated with the switched on gene is transmitted to daughter cells or even to the next generation.
Genomic Imprinting and Congenital Disease
Epigenetics plays a part in congenital disease. Genomic imprinting
means a gene is expressed differently depending on which parent it came
from. In effect epigenetics knocks out the copy of the gene inherited
from the other parent. This means that diseases that normally require
both copies of a gene to be knocked out can occur when the active copy
is damaged (the other copy having already been knocked out by genomic
imprinting).
Paramutation and Transgenerational Epigenetic Effects
In multicellular organisms, epigenetics is usually associated with
cell differentiation and the epigenetic patterns get reset in the next
generation. Some epigenetic traits can be inherited; known as
paramutation. This may help short-term adaptability by making
reversible changes in the phenotype, perhaps influenced by the
environment. Epigenetics allows genes to be expressed or suppressed
without the gene itself being mutated.
Where a mother is pregnant during famine conditions, this may have an environmental effect on the developing foetus. Genes are switched on that enable it to make better use of the reduced food supplies, for example storing any surplus calories as body fat. If the offspring grows up in non-famine conditions, those "thrift" genes are still switched on and may result in weight gain. The effects can extend to the grandchildren and influence tendencies to develop certain diseases.
Epigenetic Disease
Most people are aware of BSE (mad cow disease) which is caused by an
infectious protein called a prion. Some proteins exist in a "native"
state and also have an alternative "infectious conformational" state.
Although the genes are the same, epigenetic effects make one form
benign and the other form infectious. Prions are not always harmful; in
some fungal cells, there are advantageous prions that allow the cells
to switch on normally dormant genetic features.
Some toxins cause cancers without causing gene mutations in the affected cells. They affect the epigenetic mechanism, altering how and when genes are expressed.
Some teratogens affect the foetus by affecting the epigenetic mechanism rather than by causing a gene mutation. The effects may last throughout the individual's lifetime because epigenetic change has switched on a gene and it transcribes for products that keep it switched on after cell division. In some cases, it is possible that these epigenetic effects can be transmitted to the offspring.
DISCREDITED THEORIES OF INHERITANCE
This section mentions some of the older theories from the days before modern genetics knowledge. The role of recessive genes and mutation were not understood by most animal breeders and they relied on observation. While this allowed them to predict the likely result of mating certain individuals together, it did not explain the sometimes unexpected results that we now know are due to hidden or mutated genes (or even to paramutation). In cats, this was complicated by the fact that a female could mate with several males and each kitten in the litter could have a different father. During the early days of cat breeding, it was common practice to mate a female with 2 different males. This ensured a litter, but played havoc with deducing rules of inheritance as each male would have different hidden genes.
Mendel's theory of dominant and recessive genes was published in 1865, but the knowledge remained in the realm of scientists and was not widespread in the British livestock fancy until probably the 1930s. Even as late as
One old theory was that mating a female to 2 different males would mean the offspring inherited traits from both males - hopefully the best from each. The idea was that the kittens got (for example) a good tabby pattern from one male and the desired colour from the other male. It was not understood that only one sperm could fertilise an egg.
It used to be believed that mating a black cat to a white cat would result in grey cats. Mating grey cats to white cats was believed to give paler grey cats. This theory is called blending. The existence of dominant and recessive traits was not widely understood and breeders had no other explanation for where the grey kittens came from (white masks all other colours; black masks grey - the grey kittens occur becasue of recessive grey genes). Where an animal inherits 2 different co-dominant genes, it is intermediate in type between the parents (Tonkinese pattern). Another example where a form of blending occurs is with multiple genes (polygenes) where the overall intensity of colour depends on multiple genes inherited from the parents.
LAMARCKISM (INHERITANCE OF ACQUIRED TRAITS)
Lamarck believed that traits acquired during an individual's lifetime could be inherited by the offspring. This was sometimes used to explain the Manx cat's taillessness. According the Lamarckism, a cat whose tail had been cut off would pass on the trait of taillessness to the kittens. Before its tail was removed it would have tailed kittens. After the tail was removed it would have tailless kittens. A modern instance was that of Josephine, founding female of the Ragdoll breed. Before being involved in a car accident, Josephine was fairly wild and her kittens took after her in temperament. After being nursed back to health she was tamer and her kittens more relaxed. Did a car accident change her genetically? In reality, being nursed would have helped re-socialise Josephine so she would have brought up later litters closer to humans. Her litters were sired by various free-roaming males and the kittens also inherited different personality traits.
According to this theory, the unborn offspring were affected by the mother's environment. The female could, therefore, be mated to a mediocre stud, but if the pregnant female was then housed in close proximity to outstanding examples of her own species, the sight of those animals would cause her to impress their characteristics onto her unborn young. On the other hand, if the pregnant mother was exposed to a bad example, that would be transmitted to her offspring. This led to some unusual advice on mating and housing cats.
There is a grain of truth in the concept, but not for the reasons believed in the 1800s. Environmental factors in the womb and during life switch some genes on or off. For example, a poorly nourished pregnant female may have offspring that tend to put on weight. Poor nutrition in the womb has switched on metabolism genes that make best use of what nature assumes is a scarce food supply and the offspring may be better able to store food as fat. If the offspring is well nourished when it is pregnant, those genes will remain switched off in its own offspring.
PATERNAL IMPRESSION (TELEGONY)
The theory of telegony claimed that a female's first ever mate had a permanent effect on her and would affect all of her future offspring, even those fathered by different males. If her first ever mate was an outstanding stud, that stud's characteristics would supposedly turn up in later litters fathered by other studs. Breeders claimed to discern this influence, whether desirable or not, demonstrating the power of belief! Unfortunately, paternal impression meant that if an female animal was mis-mated to a poor quality on moggy male she was "ruined for life" and would always produce tainted poor quality offspring. Many such females were needlessly destroyed.
IMPRESSION OF ACQUIRED PATERNAL/MATERNAL TRAITS
Sometimes Paternal Impression is distinguished from Telegony and defined more in line with Lamarckism, being the inheritance of traits acquired by the sire during the sire's lifetime. For example, if the male lost an eye through injury, his offspring are likely to be born with one eye or have one eye blind or deformed. Likewise Maternal Impression could be defined as inheritance of traits acquired during the mother's lifetime rather than environmental factors impressed upon her unborn kittens.