GENETICS OF COLOUR AND CONFORMATION
This page is a simplified version of cat genetics based on Mendelian inheritance where each trait is given a symbol and is dominant, recessive or co-dominant. In cats, some traits are monogenic i.e. controlled by one pair of genes that act like an on/off switch. Other traits are controlled by polygenes (groups of genes), but because we are interested in the phenotype (visible trait) we can use a symbol and make predictions as though they are single genes. These different genes combine and interact to give a huge variety of colours and conformations. There resources on the web that can predict the outcome of certain matings (recommended if you are not a genetics ace), or that can teach you how to predict the offspring using punnet squares.
Remember, in cat breeding we are usually interested in visible traits!
The position of a gene in its chromosome is called its locus. The cat has 19 pairs of chromosome. The two chromosomes in a pair are similar (one coming from the mother and the matching chromosome coming from its father). This means 2 copies (alleles) of each gene each at the same locus of its chromosome. These alleles might be identical A-A or different A-B. To greatly simplify the process, each parent passes on ONE of those alleles to its offspring where it matches up with the equivalent allele from the other parent. This can produce different combinations, for example: A-A, A-B, B-A, B-B
Okay, so we have two alleles controlling one trait. Most likely one is dominant (is visible expressed) and one is recessive (is hidden, but can be passed on to offspring). A few genes are co-dominant and the visible effect is halfway between them. A recessive trait only shows up visibly if the cat inherits 2 recessive alleles for the trait.
BASIC COLOUR GENES
There are two types of colour pigment - eumelanin (black/brown pigment) and phaeomelanin (yellow/red pigment). There are several mutant forms of the black/brown pigment. The coat colour depends on which pigments are present. A number of other genes modify the amount of pigment in the hair shaft and whether it is laid down in alternating light and dark bands along the shaft or as a single, solid band stretching from follicle to hair tip. At the foot of this page there is a more detailed description of how melanin works to give a wide array of colours.
|
|
Dominant |
Recessive(s) |
Notes |
|
Colour/Albino |
C - full colour |
c - pink-eyed albino |
cscb = Tonkinese |
|
Agouti |
A - agouti |
a - non-agouti |
Wild-type is agouti |
|
Agouti - Charcoal Pattern |
a + APb = Charcoal Pattern |
A2 = Agouti variant found in Bengal and Savannah cats. The phenotypic effect of A2 has not been established. |
|
|
Black/brown |
B - black/brown |
b - chocolate |
Order of dominance is B > b > b1 |
|
Dilution |
D - dense |
d - dilute |
|
|
Orange |
O |
o - wild-type |
Sex-linked |
|
White Masking / Spotting |
W - White masking (masks all other colours) |
ws - piebald/white spotting |
Sb and Sp are posited modifiers. |
|
|
Dominant |
Recessive |
Notes |
|
Amber (Extension Gene/Black Modifier) |
E - black pigment produced |
e -red pigment produced |
ee genotype produces reddish pigment instead of black or blue |
|
Dilute Modifier |
Dm - modifies dilute colours |
dm - no effect on colour |
Dominant, but only affects cats that are homozygous for dilute "d" |
|
Pink Eye Dilution |
?? |
?? |
Recessive? Turns Black into a light brown colour, eyes are golden with red pupil. |
|
Colour Inhibitor |
I - gives silver undercoat |
i - no silver undercoat |
Dominant gene giving silver undercoat, works in conjunction with Wideband (Wb) in tipped, shaded and smoke phenotypes |
|
Sunshine (Siberian Golden) |
sg - gives silver undercoat |
Recessive gene in Siberian (possibly also Kurilian) breed currently only seen in agouti (tabby patterned) Siberian cats where it produces reddish areas in the black, blue etc coat. Because Sunshine is on a different locus to the Inhibitor (silver) gene, it is possible for both genes to occur in the same cat resulting in a "bi-metal" appearance where both silver and golden areas are present. |
EUMELANISTIC COLORS WITHOUT MALTESE DILUTION (BLACK, CHOCOLATE, CINNAMON). DENSE COLOURS.
The B gene is responsible for black pigmentation of the coat. It causes the production of a pigment called eumelanin (spherical particles). Eumelanin production is affected by temperature cooler temperature means more is produced, while warmer temperature means less is produced. This is why many long-haired blacks have a paler region at root of the hair this part is closest to the body and is warmer. It is also why colour tends to be denser at the points (face, ears, legs, tail) as these are cooler than the torso.
The B gene has a recessive b allele. This deforms the eumelanin into oval particles which produce the chocolate colour. Another allele, b1, is recessive to both B and b. It produces cinnamon. A black cat can be Bb or Bb1. A chocolate cat can be bb or bb1. But a cinnamon cat can only be b1b1. These are called dense colours.
DILUTED EUMELANIN COLORS (BLUE, LILAC, FAWN).
Black, chocolate and cinnamon also exist in diluted forms called, respectively, blue, lilac and fawn. These have the same order of dominance as the dense colours. They are caused by a different gene i.e. a different locus - represented by D (recessive form is d). This gene changes the concentration of pigment particles in the hair to give a paler colour. When cat inherits d from both parents, this combines with either black, chocolate or cinnamon they produce the dilute colours. Otherwise (DD or Dd) the cat has the dense colour.
The kitten of a blue cat mated to a chocolate may be black even though black is dominant over blue and dominant over chocolate. This is because the chocolate gene is at one locus and the dilution gene is at a different locus. The black offspring inherits the dense colour gene from the chocolate parent and the dominant eumelanin gene from the blue parent, but because it gets only one copy of the recessive dilution gene from the blue parent, its colour is not diluted.
|
Black/Brown Gene |
Dilution |
Dilute Modifier |
Eumelanistic Colour |
|
B- |
D- |
any |
Brown/Black |
|
B- |
dd |
dmdm |
Blue |
|
B- |
dd |
Dm- |
Caramel (blue based caramel) |
|
bb or bbl |
D- |
any |
Chocolate |
|
bb or bbl |
dd |
dmdm |
Lilac |
|
bb or bbl |
dd |
Dm- |
Taupe (lilac based caramel) |
|
blbl |
D- |
any |
Cinnamon |
|
blbl |
dd |
dmdm |
Fawn |
|
blbl |
dd |
Dm- |
Fawn based Caramel |
PHAEOMELANISTIC COLORS (RED, CREAM) AND SEX-LINKAGE (TORTIE, BLUE-CREAM).
Mendel's laws of genetics deal with either/or situations on the autosomes. There is another pair of chromosomes called the sex chromosomes and these behave a bit differently from autosomes. Sex chromosomes are X and Y. A normal female is XX. A normal male is XY. Think of the Y chromosome as being like an X that has lost a leg, and consequently lost the genes that are on that leg. This means some genes inherited from a female do not have a corresponding gene from the male. In genetic notation, this is shown as - meaning a blank space where the gene is inherited from one parent only.
Sex-linked red (ginger, marmalade, sandy, orange etc) is on the sex chromosomes. The variation in shades is due to multiple other genes. The dilute form is called cream. This gene changes eumelanin (black-based colours) into phaeomelanin (red-based colours). A male can be either - (eumelanin colours are produced) or O (red or cream are produced instead of eumelanin colours), but not both. A female may be - - (eumelanin colours), OO (red or cream colours) or O-.
O- is interesting because O is not dominant over -. In this case, each body cell has two X chromosomes which means both an O and a . During early embryo development, one of the X chromosomes is switched off at random. There is a 50/50 chance of the active X chromosome being O (red producing) or (not red producing). This means a cat with a patchwork of red and black fur (or cream/blue etc, as long as both colours are dense or both are dilute). If the switching off happens early on (when there are few cells), the cat has patched fur. If it occurs later (when there are more cells) it has brindled fur with fewer distinct patches.
For sex-linked traits, male kittens get the trait only from their mother because she supplies the X chromosome (the Y chromosome can only come from the father). Female kittens get it from both parents in equal proportions because they get an X chromosome from both parents.
DILUTE MODIFIER: CARAMEL AND APRICOT COLOURS.
Another colour gene is the Dilute Modifier (or double dilution) which changes the appearance of dilute colours. The colours become more pinkish and slightly metallic looking. This visual effect is due to the dominant gene Dm which is on a different locus to the dilution gene. It is therefore inherited independently of normal dilution. Dm only affects diluted colours, creating caramel from blue, lilac and fawn, and creating apricot from cream. Dm does not affect dense colours. The recessive dm allele has no effect on coat colour.
|
Red Gene |
Dilution |
Dilute Modifier |
Phaeomelanistic Colour |
|
O |
D- |
any |
Red |
|
O |
dd |
dmdm |
Cream |
|
O |
dd |
Dm- |
Apricot |
EPISTATIC GENES.
An epistatic gene is one that overrides the traits controlled by normally dominant genes. When we talk of dominance, we should really talk only about genes at the same locus e.g. black vs chocolate. For convenience, we often say black is dominant over blue, but these are actually 2 different genes at different loci. Dense colour is dominant over dilute colour, regardless of the underlying colour. Dense and dilute are epistatic they affect the expression of other genes. Sex-linked red is epistatic over eumelanin colours. When breeders talk about epistatic genes, they are most often thinking of masking genes such as solid white (which masks all other colours) or white spotting (which masks patches of colour). White masking and white spotting operate the same way regardless of the base colour.
WHITE MASKING AND WHITE SPOTTING (PIEBALD) GENE
White Masking and White Spotting are alleles of the same gene. White masking gives an entirely white coat, with pink nose leather and paw pads. One or both eyes may be blue and the cat may be deaf on the blue-eyed side(s) because the eye tissue and ear tissue are close together in the embryo. Orange eyed white cats are generally not deaf.
White spotting creates white patches on the body. Where the white spotting gene is active, the fur and underlying skin lack pigment the skin is pink and the fur is white. If the white spotting gene acts on the embryo cells that give rise to the eyes, this gives blue eyes. If it acts on the cells that create the skin around the eyes, but not on the embryonic eye cells, the eyes are orange, yellow, green etc. Sometimes one eye is blue and the other different coloured. Sometimes an odd-eyed cat is deaf on the blue-eyed side because the embryo ear tissue cells (next to the eye tissue cells) are affected.
Research has found that White Masking and White Spotting are alleles of the same gene (called KIT) and the order of dominance is W >> ws >> w >> wg.
The exact position of salmiak wsal in this sequence isn't yet determined.
The combinations are:
WW = homozygous white
Wws = white carrying white spotting
Ww = white carrying non-white
Wwsal - white carrying salmiak pattern
Wwg = white carrying white gloves
wsws = homozygous for white spotting
wsw= white spotting carrying non-white
wswg= white spotting carrying white gloves
ww= homozygous for non-white
wwg= non-white carrying white gloves
wgwg = homozygous for white gloves
(The jury is still out on other possible white spotting mutations for white gloves (Birman), white mitts (Ragdoll), white throat locket, white brisket spots and there is not yet a symbol for the Altai version of white spotting)
(In old notation, White Masking (WD - complete depigmentation of all the body) which is epistatic over all other colour genes was thought to mask a separate White Spotting gene WS. In old notation the corresponding recessive allele was wd and did not cause depigmentation. Because WD was considered epistatic over WS a genetically white cat WD/Wd) could supposedly mask white spots which could show up in non-white offspring, while, while a WD/ WD cat would always have white offspring. Older texts refer to White Spotting as S with a recessive allele s this is now ws and recessive alleles are w and wg see below.)
ws is extremely variable in how it shows up. A cat with a single copy of ws might have a small amount of white or a large amount. Most often, though, cats with two copies of ws have more white, and may even be solid white. Selective breeding can fix the amount of white because other genes, known as polygenes, can influence colour distribution e.g. Some breeds have been fixed for gloves or mitts.
BIRMAN AND RAGDOLL MITTED PATTERN GENE
Some white-marked patterns are very stable, which suggests a different white spotting gene/allele. For example the mitts and inverted V of Birmans and Ragdolls are stable patterns. If this is a different gene, then it must be homozygous recessive because it breeds true (if it were dominant, non-white-marked variants would occur). This is due to selective breeding for homozygous recessive cats that always breed true.
Mating a Birman to a non-mitted pointed cats has produced non-mitted pointed offspring. When bred together, some of their offspring (about 25%) were mitted and some were non-mitted. The gene symbol wg is Birman gloving . Some other breeders have said that non-mitted-pointed cats bred to mitted-pointed cats have produced a mix of mitted and non-mitted kittens. This could happen if the normal white spotting gene ws (previously S) is affected by a separate gene that controls where and how much white appears. The mitted gene wg (mitted) is recessive to W and ws so that all Birmans are homozygous for wg.
(Modern notation for Birman gloving is wg and replaces G or mit. sb was sometimes used, but sb was intended for the mitting on non-Birman cats. Previous theories had suggested that all Birmans were homozygous for both mit AND dominant S (the alleles being shown as SS mit mit) since cats with Ss Mit mit (heterozygous white spotting) would give non-mitted pointed cats i.e. ss mit mit offspring and this does not occur in real life.)
The Ragdoll mitted pattern closely resembles the Birman mitted pattern as the founding female of the breed may have mated with Birman males (among other suitors). However, Ragdolls have additional piebald spotting creating a white strip on the belly of mitted Ragdolls. The Ragdoll mitted gene is probably wg + modifiers (and in older articles was often shown as sm a supposedly incompletely recessive allele of white spotting).
Mitted Ragdolls have considerably more white than in Birmans long boots at the back and a white strip on the underside, from the chin to the base of the tail so there are some unidentified modifiers at play. The ideal Bicolour Ragdolls is no more than 1/3 of white on the back, white legs and the underbody almost entirely white with white extending to the face in the shape of an inverted V this stable distribution also suggests polygenes at play. The order of dominance and modifiers would also explain the high mitted, mid-high white and high-white patterns.
(If you prefer to think in old notation for convenience, ss = Colourpoint Ragdolls (without white); sm s = mitted Ragdolls; Ss = ideal bicolour Ragdolls; Sm sm = high mitted pattern; S sm = mid-high white and SS = high white pattern. Purely for prediction purposes, some breeders have adopted a notation of S1, S2 and S4 to describe the trait: ss = no white spotting; S1 S1 = Birman gloving; S2 s = Ragdoll mitted; S2 S2 = high mitted; S4 S2 = mid-high white; S4 S4 = high-white.)
PATTERN GENES
Colourpoint is not included here as it is a form of albinism. These genes affect the distribution of colour on the body. Some of the posited genes have been given symbols by breeders, but are not widely accepted. You will find that outdated texts give Ta as ticked tabby, Tm as mackerel tabby and Tc as classic tabby and describe them as alleles of a single "tabby" gene. This was later proved incorrect. Tm and Tc are no longer used; mackerel is now Mc and classic tabby is mc. Ticked tabby is a different gene and remains Ta although some early research designated it U for "unstriped tabby".
|
|
Dominant |
Recessive |
Notes |
|
Bengal Glitter |
|
gl |
|
|
Melanin Inhibitor |
I - silver |
i |
Silver (ii + Wb = gold) |
|
Ojos Azules |
?? |
|
Dominant, causes blue eyes and white splashed extremeties |
|
Spotted Tabby |
Sp |
|
Egyptian Mau type spots |
|
Tabby Pattern |
Mc- mackerel |
mc - classic |
|
|
Tabby Pattern Modifier - size |
Thl - large pattern classic tabby |
ths - small pattern classic tabby (Sokoke type) |
Posited for Australian Mist, interacts with mackerel or classic tabby to determine blotch size/density |
|
Tabby Pattern Modifier - spots |
Pmf - modified tabby |
pmu - unmodified tabby |
Posited for Australian Mist, interacts with mackerel or classic tabby to give small or large spots respectively |
|
Ticked Tabby |
T a- Abyssinian ticked |
ta - non Aby ticked |
Originally thought to be an allele of Tabby Pattern, but now known to be on a different locus |
|
White Masking / Spotting |
W - White masking (masks all other colours) |
ws - piebald/white spotting |
Sb and Sp are posited modifiers Breeds other than Birmans have white mitting, but may not have the "White Gloving" gene |
|
Wide band |
Wb |
|
Determines width of silver/gold undercoat band between hair follicle and first band of pigment. |
TABBY (AGOUTI) PATTERNS.
The classic theory of tabby (agouti) patterns is based on one Tabby locus. The modern theory involves two tabby pattern genes plus some modifiers. Each agouti hair has several colour bands laid down while the hair grows. The darker bands contain normal shaped pigment granules. The light band contains more elongated granules which look lighter and more reddish. The agouti gene, A, allows this banding to occur. The recessive form, non-agouti a , means dark pigment is produced along the whole length of the fur and suppresses the production of elongated granules. The wild type is agouti. In patterned tabby cats (e.g. classic/blotched, mackerel etc) some areas are agouti (ticked) and some areas are non-agouti (markings).
There are three main tabby patterns: ticked, mackerel and classic/blotched. All three have facial markings. The ticked pattern (e.g. Abyssinian) has no markings except for the head and some residual markings on the legs and tail. The rest of the body has agouti hairs. On the belly, where there is naturally less pigment, the agouti banding is less visible (less contrast between the bands). The mackerel pattern is the striped tabby pattern. The classic/blotched pattern has thicker, darker markings including the bull s eye markings on the flanks.
These three patterns are due to three alleles of the tabby gene. Ticked (Ta) is dominant. Mackerel (T) is dominant of blotched, but recessive to ticked. Classic (tb) is recessive. In addition there is the spotted pattern, where the lines become spots, arranged in vertical lines (like broken mackerel stripes). This appears to be due to the mackerel tabby gene + some polygenes that break up the stripes. Studies in the Australian Mist in the 1980s showed that these polygenes also act on blotched tabby. Mackerel tabby + polygenes = smaller vertically aligned spots. Blotched tabby + polygenes = larger spots that look more random because they are caused by breaking up the blotched pattern.
There is also a tabby pattern size modifier seen in the Australian Mist and some other breeds. Again, these are probably polygenes, but they are simplified as a single gene symbol. Thl produces a large pattern classic tabby while th recessive form, ths, produces the small pattern classic tabby (Sokoke type pattern). Such a gene would interact with mackerel or classic tabby to determine the blotch (marble) size and density. The second posited gene (or polygene complex) is the spotted pattern modifier or Pmf (dominant form) and pmu (recessive unmodified pattern ) that gives small (Pmf + mackerel tabby) or large (Pmf + classic tabby) spots..
In breeds that carry genes from a wild ancestor (Bengal, Savannah, Safari) there are other spotted patterns due to the wild genes. Bengals may have larger, rosette shaped spots in a more random arrangement. The best rosettes can be found in lines that include Margay genes from a foundation cat imported from the defunct Bristol hybrid breed.
The marbled tabby pattern that first appeared in the American Shorthair and then in the Bengal, has very elongated, irregular bull s eyes and spirals, and thicker, often closed lines. This appears due to the blotched tabby gene tb combined with modifier genes. Alternatively there may be another tabby allele (tm similar to the suggested Pmf gene!) with partial dominance. Marbled tabby bred to spotted tabby show a breaking up of the marbled pattern into rosettes and irregular spots that are partially distributed according to the marbled design.
The wild-type agouti pattern is very close to ticked tabby and is due to the same gene, but with necklaces and tabby bars (rings) on the legs and tail. This is because it does not have the polygenes found in pedigree ticked tabbies (e.g. Abyssinian) that remove those residual markings. In eumelanin colour (black series, dense and dilute) cats the agouti gene acts on the fur and on the nose leather, turning the nose leather a shade of red/pink with nose-liner in the base colour.
If tabby pattern was due to different alleles at the same locus, with partial dominance in the order Ta > T > tb then these three patterns cannot occur in a single litter to the same parents. For example:
TaT x TaTb gives TaTa, TaT, TTb or TaTb = 75% Ticked and 25% mackerel, but no classic tabbies.
All 3 patterns can occur in the same litter. This means the ticked tabby allele ( unpatterned tabby ) must be at a different locus, and is partially epistatic to the mackerel/spotted and classic/blotched alleles that are at the usual locus.
Even the ticked pattern is variable. Abyssinians should have no bars on the limbs or tail. Singapuras should have some markings on the lower legs and tail. Many other ticked cats have bars on the torso, neck, legs and tail and facial markings. TICA recognise 2 different ticked tabby patterns: ticked (all hairs, except on the belly, are ticked, with no bars) and agouti (bars on legs and tail). The residual markings have been explained as breaking through from a recessive tabby gene (classic or mackerel). There are 2 arguments against this. Firstly, the ticked pattern becomes more dominant with age, but Singapuras retain their residual markings with age (genetically stable pattern). Secondly, the transmission probability of the agouti pattern should be 50% agouti (with residual markings), 25% purely ticked (no markings) and 25% mackerel. Purely ticked Singapuras or mackerel Singapuras do not exist. This means there must be a separate allele on the second Tabby locus, a new third Tabby locus, or a group of polygenes affecting the residual pattern. For prediction purposes, it is easier to assume 2 Tabby loci plus some polygenes to explain minor variations.
Ta (ticked tabby) is at one Tabby locus. T (mackerel) and Tb (classic/blotched) are at a different Tabby locus. The gene that removes the residual markings is termed U (Unstriped/Unpatterned) and it has a recessive allele, u, at the same locus. The recessive form does not mute the pattern and it s easier to refer to it as recessive ta.
Ta t- Ticked pattern, partially or totally masking mackerel-spotted
Ta- tbtb Ticked pattern, partially or totally masking classic
tata tt Mackerel or spotted pattern
tata ttb Mackerel or spotted pattern (partially) dominant over classic
tata tbtb Classic pattern (or marbled: tata tmtm)
How The Ticked Genes Masks The Mackerel Or Classic Patterns
In the single locus theory, the ticked allele was partially dominant over mackerel/spotted and classic. The recessive pattern was visible at birth, but faded in adulthood. A homozygotous ticked (TaTa) would be completely ticked at birth. But with the two loci theory the other patterns are present (on the other locus), but masked. In practice, even if a cat is homozygous TaTa some cats show a breakthrough of the tabby pattern at the other locus. Other cats don t show any residual or ghost markings. This leads us to polygenes that modify the pattern.
Polygenes for agouti band distribution: band frequency and wide-band genes, as for instance in the shadeds
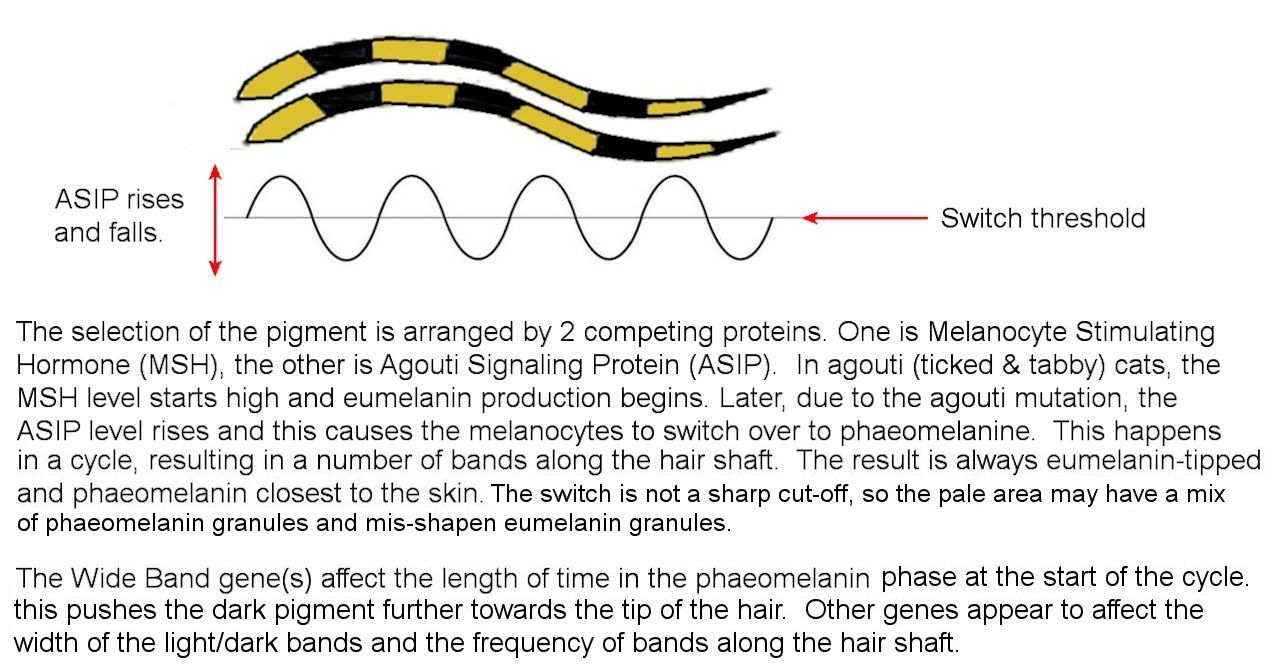
The production of melanin results from the melanocyte stimulating hormone (MSH) binding to the melanocortin receptor (MC1R). MC1R is coded by genes at the extension locus (we ll look at this locus later on). The Agouti gene (and its wide-band modifiers) codes for Agouti Signalling Protein (ASP) that binds to MC1R and blocks MSH. If this occurs periodically, it causes alternating bands of black-based and red-based pigment. If it occurs permanently, you get red or cream colour hair. The level of ASP rises and falls. The hair tip is always eumelanistic, and the rest of the hair has alternating bands. The frequency of these bands is related to the time needed for ASP levels to cycle up and down. This cycle is irregular, hence the bands are not uniform width. Genes such as Wide band affect the cycle length. There are other band frequency genes at different loci.
The masking effect of the ticked pattern relates to the frequency of banding. If adjacent follicles are well synchronized and the hairs have same size bands at the same places (high coherence) the ticking appears more uniform and looks darker. In Abyssinians, there is a largely uniform ticked area which ends in a distinct line at the belly. In shaded cats, it is the non-uniform frequency of agouti bands produces the sparkling effect of the top colour. These traits are inherited so there must be one or more genes controlling band frequency. Unlike single-gene traits, perfecting a pattern controlled by polygenes can take many generations of selecting the best marked cats for breeding.
One gene was originally termed Confusion, but is now known as Frequency, F (strongly expressed, e.g. in a perfect classic tabby pattern with no agouti hairs breaking up the black markings, it is termed FF or full coherence). Dominant F gives high coherence of band frequency distribution (as in Abyssinians); recessive f creates a disordered pattern (as in shaded silvers) and masks a striped, spotted or classic pattern.
Polygenes For Agouti Hair Distribution: Area Coherence
In a perfect tabby, there is a sharply defined boundary between the markings and the agouti area. In a less perfect classic tabby this boundary is blurred by agouti hairs. In agouti hairs, there is a pale band (silver in silvers, pale brown in non-silvers) at the base of the hairs, close to the skin. These base bands are not visible unless you part the coat and they do not disrupt the pattern. Agouti hairs with many bands are visible and they mute the tabby pattern. Bengal breeders refer to this disruption as ticking, by which they mean the presence of banded agouti areas inside the black spots. The intrusion of agouti hairs inside the markings can be desirable to obtain a perfect shaded silver because it disrupts or removes the remaining stripes in the torso but they must also low coherence to do this.
How is coherence inherited? It appears to be controlled by polygenes, but for simplicity breeders represent it as a simple Mendelian gene termed Agouti Distribution, Ad (also known as Chaos) as this helps them predict the outcomes of matings. Dominant Ad produces a lack of coherence (or a lot of disorder/chaos), while the recessive ad allows order i.e. the markings are not muted. The Area Distribution locus controls the erasing of stripes from the torso. Leg and tail bars, and also necklaces, are harder to erase and probably controlled by genes at yet another locus.
GRIZZLED TABBY
The grizzled pattern appears in the Chausie breed, probably due to a version of the tabby gene inherited from its wild Jungle cat ancestor. Because Chausies are only bled in eumelanin (black) colours, the grizzled pattern shows up as fully coloured legs and face, including black nose-leather (unlike other tabbies). The hairs on the body have up to five agouti bands that have the pale yellow color, but with cool tones, so they look like silver tipped hairs. It is not related to the silver (inhibitor) genes found in domestic cats.
So far is isn t known if this pattern is due to an imported dominant allele at the tabby locus (a grizzled tabby allele), or if it is due to an epistatic gene at a different locus. If it is an epistatic gene, it may be on the Extension locus E, as seen in steel colour rabbits. The epistatic gene theory is preferred because the nose leather in other tabby cats is reddish; but in grizzled Chausies and non-agouti cats it is the same colour as the based colour. Regardless of whether it is a tabby allele or an extension gene, the grizzled pattern allele is dominant over all the other tabby patterns.
Because the tabby pattern is controlled by genes at one locus, while the agouti/non-agouti (or non-patterned/patterned) trait is at a different locus, solid colour cats still have an underlying tabby pattern it just doesn t show up except under certain lighting conditions!
The tabby alleles are not completely dominant over each other. A cat with both Ta and Tb (ticked partially dominant to blotched) may have more marking on the legs, tail and face while the body ticking is a bit blotchy. A cat with both Ta and TM may look pinstriped, while one with Ta and Tm + spotted polygenes can look freckled.
RED TABBIES
The inter-action of tabby genes on red/cream tabbies is the same, but the visual effects are a little different. The non-agouti gene is much less effective on red pigment (phaeomelanin colours) than on black-based (eumelanin colors). As a result, the tabby pattern cannot be completely cancelled out by non-agouti. Breeders have achieved cats that look nearly solid red by careful breeding that has brought together various polygenes that, by their cumulative action, have reduced the contrast between background and foreground colours.
To compare how the non-agouti works on red and black pigments, look at a tortie cat (with or without white) and you will see solid black patches but the red patches have residual markings (so the normal black/red tortie is really black, red and cream). In dilute torties this will be solid blue plus two shades of cream.
Unlike eumelanin colour cats there is no sharply defined tabby pattern on / tabby pattern off switch. In non-agouti red cats the solidness is a matter of degrees depending on what polygenes are in the mix.
In agouti (patterned tabby) tortie cats there are tabby patches of eumelanin colour and tabby patches of phaemelanin colour. These are tortoiseshell tabbies (torbies).
THE SILVER SERIES (SMOKE, SHADED, SILVER TABBY).
The silver series are smoke, silver tabby, shaded silver and chinchilla/tipped. The red-series (phaeomelanin) equivalents are often called cameos because the pinkish colour resembles cameo jewellery. In silver cats, the coat is depigmented at the base i.e. it is silver white without any rufousing (reddish tones). In silver tabbies, the agouti parts of the coat are silver, but the marking colour is solid from base to tip of the hair. A good chinchilla should have no trace of tabby markings, with only the hair tips being pigmented (tipped). Shaded silvers are more heavily tipped (about one third of the hair s length). Smokes have the greatest degree of pigment about one half to one third of the hair is coloured. A smoke is a non-agouti cat while the others are tabby patterned cats with varying degrees of colour and pattern.
|
SOME OLD THEORIES FOR SILVER & SMOKE AND WHY THEY DON'T WORK
1 Silver = Single Inhibitor Gene At first, silver was attributed to a single inhibitor gene, I that inhibited pigment production and cancelled rufousing (the reddish colour shades of eumelanistic cats, particularly in the agouti areas of brown tabbies). Shaded silvers and chinchillas both have ticked patterns. The addition of the I gene turns the hair colour into silver at its base and cancels all reddish tones from the lighter bands, making them silvery white. If the I gene acts strongly it produces chinchillas. If it acts mildly it gives shaded silvers. Silver tabbies are agouti with blotched or mackerel/spotted pattern. The I gene acts best on the agouti areas, turning them silver. Because ticked tabby cats may also have a recessive blotched or recessive mackerel tabby allele, there may be a faintly visible tabby pattern e.g. on the face and forehead. These all reddish nose leather with dark nose liner indicative of an agouti cat. We know the I gene works on non-agouti fur to give smoke, so why does it leave the non-agouti markings of a tabby patterned coat untouched? Why is the fur in the tabby markings solid to the base (at least in good quality silver tabbies, there are some intermediates that show a smoky tabby pattern)? 2. Silver = Silver Bleaching Gene + Eraser Gene (Flawed Theory) Another theory tries to answer that by suggesting two interacting genes called silver bleaching (Sv) to cancel rufousing, and eraser (I) to cause silver at the base of the hair. The posited Sv gene cancels rufousing in eumelanin colours, but not so much in phaeomelanin colours - leaving behind warmer tones. Yet another theory gives two forms of I- Sh (shaded silver) is the mild form, while Ch (chinchilla) is the strong form. However there are heavily marked Chinchillas and lightly marked shaded silvers that sit midway between the two patterns i.e. silvering is a gradual effect, not an either/or effect. This suggests polygenes affect the degree of silvering/bleaching in the agouti fur. The lack of rufousing makes the black marking look more intense. According to this theory, polygenes could explain the existence of smokey tabbies. With only the silver gene, but no eraser gene, shaded and chinchilla cats would have a coloured band near the skin, something not seen in cats. In smokes (non-agouti silvers), the silver gene on its own would give cool-toned (unrufoused) solid colour cats, but silver + eraser would give the smoke pattern. If smoke cats heterozygous for both silver and eraser (sv and i) were bred together, they should produce some solid colour offspring (svsv ii). If shaded or chinchilla cats heterozygous for both silver and eraser (sv and i) were bred together, they should produce some full colour ticked tabby offspring (svsv ii), albeit less rufoused than an Abyssinian. Shaded silvers and chinchillas are selectively bred using cats that lack rufousing polygenes or have unrufousing polygenes (we don't know exactly what the polygenes do!). Cats that are svsv Ii or svsv II (non-silver, but plus eraser) are pale ticked tabbies, but without the cool tones caused by dominant Sv They would have a golden base colour shade rather than a reddish one. These are shaded goldens and they are only recognised in eumelanin (black-based) colours not in red-based colours. Red-based versions are visually similar to red or cream ticked tabbies. Likewise, golden tabbies would result from breeding together silver tabbies that carry the recessive sv allele. A golden tabby (svsv) with dominant I would be golden tabby shell or shaded depending on the amount of pigment present. A golden tabby with only recessive i would not have depigmented hair roots. In theory, a non-agouti cat with I- svsv should be a golden smok, but golden smokes don't exist in reality. This means the I + Sv theory is also flawed. There are 2 arguments against I and Sv genes: the non-existence of shaded cats with a dark band at the base of the fur, and the non-existence of shaded goldens. Maybe Sv/sv cannot activate unless dominant I is also present. This is similar to the epistatic golden modifier theory. 3. Golden Epistatic Gene According to the epistatic golden modifier theory, goldens are caused by an epistatic gene, g, that affects the expression of gene I. This is a bit like the way dilution influences the expression of black, chocolate and cinnamon. The other allele, G, has no action i.e. it does not produce golden pigmentation at the base of the hair. This posited gene symbol is not used; it would clash with G for Birman Gloving. 4. Golden Phenotype = Brown Ticked Tabby Another theory identifies shaded goldens as ticked tabbies. Because pigmentation is heat-dependent, the coat colour is usually lighter near the skin. Therefore the golden and the ticked tabby phenotypes are similar. However . . . this does not explain why the goldens are paler near the skin than the normal ticked tabbies. This brings us to polygenes and the Wide Band effect. Goldens are bred from shaded silver and chinchilla lines, and those cats are selected for the greatest amount of depigmentation (greatest amount of wide banding ) near the skin. Two chinchillas/shaded silvers heterozygous for Inhibitor can produce ticked tabby offspring with various degrees of Wide Banding. These range from cool toned ticked tabbies (because breeders have also bred out the rufousing polygenes) to chinchilla/shell/shaded goldens. |
5. Inhibitor + Wide Band
This two-gene theory best explains the existence of silver tabbies with markings colored all the way to the root as well as silver tabbies with a depigmented zone near the skin. So the currently accepted theory is Inhibitor + Wide Band (Wb). Although treated as a dominant gene, the variable expression suggests polygenes. By selectively breeding the palest chinchillas, breeders have brought together multiple polygenes.
According to this theory, the Inhibitor (silver) gene cancels rufousing. An additional dominant gene (more like a set of polygenes) called Wide band (Wb) makes the agouti bands wider. This explains the non-existence of golden smokes, because Wide Band only acts on agouti fur. Chinchilla = ticked tabby = Inhibitor + Wide Band. Golden = brown ticked tabby + Wide Band. It doesn t explain why silver smokes have a depigmented silvery band, perhaps Inhibitor does that on its own.
Cats registered as goldens have varying levels of depigmentation, ranging from no depigmentation through to no top-colour. This continuum points to polygenes instead of a single Wide Band on/off gene.
What if the shaded silvers used to produce goldens are heterozygous for ticked tabby gene i.e ticked carrying mackerel, or ticked carrying classic? In silvers these are usually shaded silver and may show faint tabby markings on the legs. In goldens, the kittens carrying the mackerel or classic gene have visible stripes on the legs. The Wide Band polygenes transform spotted, mackerel and classic tabbies into golden spotted, golden mackerel and golden classic tabbies. The normally brownish agouti background colour becomes a warm apricot colour.
This Inhibitor + Wide band theory means no need for an eraser gene (which would permit golden smokes). As for the inconvenient issue of some silver tabbies being depigmented at the base of the hair while others are pigmented, most likely this is also down to polygenes and, for convenience, include under the umbrella term of Wide Band.
Because the effect of Wide Band is less dramatic than the effect of Inhibitor, the golden tabby colours are harder to identify, especially in kittens where there might be ghost patterns due to the presence of a recessive tabby pattern gene. Because Wide Band produces a reddish colour, it is hard to identify red/cream tipped goldens or red/cream shaded goldens.
6. Ultra Wide Band Polygenes
Breeders have selected for the palest possible chinchillas which means they have brought together more and more of the polygenes to lighten the hair shaft and widen the depigmented bands. Traditionally, chinchillas have a sparkling effect caused by the coloured fur tips. However, some breeders have taken the Wide Banding to such extremes that the top-colour is no longer visible at the hair tips (at least not to the naked eye). The coloured tip is too small to accommodate many pigment granules. This produces chinchillas that appear solid white, but have dark eye liner, dark paw-pads and greenish eyes. The dark nose-liner may be lost due to breeding for extreme Side Banding.
When these chinchillas produce goldens, the effect of so many polygenes is even more dramatic. The back and sides of the cat look solid red-gold. This extends along the legs and tail and onto the forehead, nose and cheeks. However the underside appears almost pure white including white toes, white throat and white chin. Once again, the pigmented tips are so small that we just cannot see the dark pigment. Often, the only place the top-colour is visible is the end of the tail. This distribution of colour is the same as in exhibition quality ticked tabbies (look at a blue Abyssinian for an example). The presence of Wide Band removes almost all dark pigment to exaggerate that colour distribution. This is particularly seen in British Shorthairs bred in Russia and parts of Germany and some breeders refer to this phenotype as copper to keep it separate from normal tipped goldens.
For convenience, we can denote this effect Wb, Wb+ and Wb++ i.e. normal Wide Band, Wider Band and Extreme Wide Band.
Eye Color Of Shaded Silvers And Goldens
The usual eye colour in ticked tabbies is copper (Persians) or gold, or hazel or green (Abyssinians). Up until the late 1890s, copper eyes were standard for eumelanistic silvers. Green-eyed or blue-green-eyed cats were originally disqualified, but later became the standard. Eye colour is copper or gold in the phaeomelanisitic silvers. Copper eyes have been re-introduced into eumelanistic silvers resulting in the pewter. This demonstrates that eye colour is not affected by the silver Inhibitor gene.
Eye colour is mostly controlled by a dominant/recessive pair of alleles + some polygenes that influence the shade of the basic colour. Black-based shaded silvers with copper eyes are far less common than their green-eyed counterparts which suggests some sort of link between coat colour and eye colour in cats with the Inhibitor gene. It is possible that the Inhibitor gene is located close (on the chromosome) to the gene for emerald eye colour so they tend to be inherited together (alternatively neither is inherited). This is known as persistence. But because early silvers were copper-eyed, it is equally likely that selective breeding has removed the copper eye colour from the gene pool.
THE EXTENSION LOCUS - AMBER COLOURS.
The coat (and skin) pigmentation is due to variants of the melanin pigment, either eumelanin (black-based pigments) or phaeomelanin (red-based pigments). These are produced by melanocytes. First the difficult bit why do pigment cells produce either black or red pigment?
The production of melanin results from the melanocyte stimulating hormone (MSH) binding to the melanocortin receptor (MC1R). MC1R is coded by genes at the extension locus. When binding occurs, the melanocyte produces eumelanin. If this was all there was to it, we would have no red-based colours or banded fairs. The sex-linked Orange gene permanently stops production of eumelanin and allows production of phaeomelanin. The Agouti gene causes production to switch between eumelanin to phaeomelanin to produce bands. White masking and white spotting genes stop pigment production.
The Agouti gene (and its wide-band modifiers) codes for Agouti Signalling Protein (ASP) that binds to MC1R and blocks MSH. If this occurs periodically, it causes alternating bands of black-based and red-based pigment. If it occurs permanently, you get red or cream colour hair. The production of eumelanin instead of phaeomelanin is also regulated by the enzyme tyrosinase. If the level of tyrosinase decreases in the melanocyte, phaeomelanin is produced. The E/e genes at the extension locus control the level of tyrosinase in the melanocyte. The action of the Extension gene is independent of the Agouti locus, and can may appear in both agouti and non-agouti cats.
The dominant E (EE or Ee) results in high levels of tyrosinase, therefore black pigment. If the cat is homozygous for recessive e (ee) there will be low levels of tyrosinase and red pigment will be produced. . In agouti cats with black-based colours the hairs have alternating bands of dark pigment and brown/reddish pigment. In agouti cats with red-based colours, the alternating bands are red and an orangey colour. But where the cat is ee homozygous for non-extension it produces red pigment despite being genetically black-based, which results in Amber (Norwegian Forest Cats) and Russet (Burmese). A somatic mutation of the extension gene might be responsible for anomalous reddish patches on some male cats.
There may be other alleles of the Extension gene not yet identified in cats. For example, an Extension gene allele in rabbits produces unpigmented ( silver ) hair tips. Other alleles change the distribution of red and black pigments on the body (e.g. bay horses with a reddish body and black legs). In rabbits there are at least 4 variants of the gene (alleles). In order from dominant to recessive in rabbits, and with feline terminology, these are:
1: Steel, Extension of Dark Colour (Es) a nominee for grizzled colour on black-based colours (Chausie), or possibly pseudo-cinnamons and blue-gingers (from red-based colours temperature sensitivity would explain why the bluish pigment is usually found on the lower legs, muzzle and end of tail)
2: Full Colour Extension (E) normal agouti banding
3: Harlequin (Japanese Brindle) Pattern (ej) non-sex-linked tortoiseshell pattern
4: Extension of Light Colour/Non-Extension of Dark (e) russet/amber
Breeders have only recently taken notice of colours produced by the Extension gene. Several Norwegian Forest Cats change their colours from black or blue to a reddish or cinnamon colour as they matured. Their paw pads remained black or slate gray. This occurred in tabbies and in solid cats and also in cats with the silver Inhibitor gene. Cinnamon and chocolate do not exist in Norwegian Forest Cats. The two new colours were called amber (the dense colour) and light amber (dilute version).
Grizzle - Tabby Gene Or Extension Gene?
The Grizzle colour/pattern was imported from Felis chaus into the Chausie breed. The Chausie is only bred in black-based colour. It might be a new dominant allele at the Tabby locus. Grizzled hairs are alternately banded with red and black pigment like agouti hairs. This visually resembles the steel rabbit with its silver-tipped fur, but unlike steel the hairs are banded. Experimental matings between Grizzled and Amber cat would allow breeders to discover if it is an Extension gene allele, but from the breeders viewpoints, this is an undesirable mating and could pollute both breeds.
Non-Sex-Linked Tortoiseshell?
In cats only E and e have been documented so far, but in rabbits there are at least 4 variants of the Extension gene (alleles). In order from dominant to recessive in rabbits, these are:
1: Steel, Extension of Dark Colour (Es)
2: Full Colour Extension (E) normal agouti banding
3: Harlequin (Japanese Brindle) Pattern (ej) non-sex-linked tortoiseshell pattern
4: Extension of Light Colour/Non-Extension of Dark (e) russet/amber in cats
The "ej" (Japanese brindle) allele creates Harlequin in rabbits. Very early in the development of the embryo, Harlequin causes the switching mechanism to break completely. Some cells lose the ability to make black pigments, while others are stuck permanently in black mode . The cells continue to divide and they form clonal patches of either red or black. Being recessive, the presence of dominant E (Eej) gives solid black (it overrides the action of ej). In rabbits, the presence of recessive e (eje) gives wholly red colour. But ejej (homozygous ej) gives a mixture of red patches and black patches. In cats, this would probably be indistinguishable from a normal tortoiseshell pattern because regardless of the mechanism - the visual effect is a turning on of either red or black pigment. The major difference would be the mode of inheritance ej isn t sex-linked and it could be transmitted and displayed by both males and females.
Black and Tan?
In rabbits, the black-and-tan pattern is produced by a recessive variant of the Agouti gene called at. This version of agouti hasn t been found in domestic cats. The version acts so that wherever a region normally starts black (e.g., black tipping on agouti hairs) the gene keeps the extension gene ( E ) n 'black' mode. Meanwhile, wherever the hairs don t have black tipping (e.g., belly, inside ears, under jaw) the extension gene sticks in red mode .
POINT COLOURS: SEPIA (BURMESE COLOUR RESTRICTION) MINK (TONKINESE COLOUR RESTRICTION) AND COLOURPOINT (SIAMESE COLOUR RESTRICTION).
The albino series of genes act on colour distribution and intensity. In order of dominance, the alleles are C (full-colour), cb (sepia), cs (colourpoint almost equally dominant with sepia), ca (blue-eyed albino) and c (pink-eyed albino). The sepia allele is only partially dominant over the colourpoint allele and if a cat inherits one of each it will have an intermediate mink colour as found in the Tonkinese.
The cb allele makes pigment cells longer, and the resulting lighter colour has reddish tinges. Because the effect is temperature related, the points (head, ears, legs, tail) are slightly darker than the torso. should be of the same color shade of the body, but since pigmentation is temperature related, the points are often slightly darker. Note: instead of black sepia the colour is called seal or sable. The Singapura is genetically a sable ticked tabby. cb also results in gold (preferred in Burmese), green-gold or yellow-gold eye colour, but never in copper or green eyes.
cs produces the Siamese-style pointed colours, known as the Himalayan pattern (from rabbits). The body is light coloured and the points are much darker (with a reddish hue), giving a higher contrast than the sepia colour restriction. As a cat ages, the body temperature becomes cooler and therefore darkens. A genetically black colourpoint is seal at the points, not black. cs also causes blue eye colour.
cbcs gives an intermediate colour called mink where the points are intensely coloured and somewhat reddish while the body is a paler version of that colour. This is paler and has a higher contrast than sepia, but darker and lower contrast than colourpoint. The eye colour is also intermediate between green and blue.
Mocha, cm, appears to be another gene in this series and possible co-dominant with colourpoint. It has not been fully investigated.
ca and c are albino. ca stops pigment production on both body and points, resulting in a white cat with pale blue eyes. c takes this one step further and the eyes are transparent with a pink hue due to the blood vases in the retina. Due to the structure of the lens, a true pink hue is rare because light refraction produces a violet tint.
AMERICAN OJOS AZULES AND RUSSIAN TOPAZ GENES.
Blue eyes can also result from other genes than White Masking, Colourpoint or Albino. The Ojos Azules gene, Oa, produces blue eyes and a white tail tip. However it also has potentially devastating effects on embryo development and is no longer bred. A more recently discovered gene (no symbol yet) is Russian Topaz (sometimes called Russian Ojos Azules) which is not related to the American Ojos Azules. This seems to be free of the health issues. Some cats, presumably homozygotes, have eyes so dark that they appear black.
PINK-EYED DILUTION (UKRAINE CHOCOLATE)
Pink-eyed dilution was lost in cats in 1961, but re-emerged around 2009 and is being bred in the Donskoy. It changes black to a light brown colour, which resembles chocolate, and the eyes have a red pupil and gold flecked red/pink irides (the coloured section around the pupil). Genealogical data confirms it to be an autosomal recessive. The gene locus has not yet been identified or given a symbol. Interactions with genes other than B (black, dense) are not known.
COAT STRUCTURE:
HAIR LENGTH
Coat length is primarily controlled by the Length (L) gene. Dominant L gives a short-haired cat. Recessive l produces long hair. Various modifier genes affect these so the coat length can range from the short, sleek Oriental coat, through semi-longhair and the other extreme of the long-haired exhibition Persian. Because L (short-hair) is dominant, two Exotic Shorthairs can produce long-haired offspring. Because the Exotic Shorthairs have also been bred for a particular set of modifier genes, their long-haired offspring probably won t have the same coat quality as Persians bred from generations of Persians. This is why they are often registered as Exotic Longhairs or Exotic variants and are not registered as Persians.
|
|
Gene Symbol |
Notes |
|
Cornish Rex |
r |
recessive |
|
Devon Rex |
re |
recessive |
|
Dutch Rex |
Rd |
dominant |
|
German Rex |
gr |
recessive |
|
Long hair |
l |
recessive |
|
Hairless (French) |
h |
recessive |
|
Hairless (Hawaiian) |
hr |
DNA analysis found this to be same mutation as Sphynx with other effects being due to interaction/recombination of genes on other loci |
|
Hairless (Redcar hairless) |
hd |
Recessive, lethal (British hairless mutation) |
|
Hairless (Mexican) |
?? |
Never identified, research in newspapers has found that this term was a generic name for early hairless cats regardless of parentage or geographical origin. |
|
Hairless (Peterbald & Donskoy) |
Hrbd |
dominant (incomplete penetrance) Combinations: Hrbd > Hr >hr |
|
Hairless (Sphynx) |
hr |
Recessive; Canadian Sphynx breed. |
|
LaPerm |
Lp |
dominant |
|
Oregon Rex |
ro |
recessive |
|
Recessive Shorthair |
?? |
recessive. Shorthair is normally dominant over longhair, but some Persian lines (should breed true for recessive longhair gene) have produced shorthair offspring. |
|
Selkirk Rex |
Se |
dominant (the symbol Rs has also been used) |
|
Wirehair |
Wh |
Dominant (poss incomplete) |
|
York Choc Undercoat |
Yuc |
Dominant, lacks undercoat (posited) |
RUSSIAN HAIRLESS TEST MATING EXAMPLE
Russian Hairless (Donskoy or Peterbald) is HrbdHrbd haha and has 1 possible gamete (Hrbd ha) crossed to a Normal Haired Cat is HrHr hiha and has 2 possible gametes (Hrha and Hrhi).
|
|
Hr ha |
Hr hi |
|
Hrbd ha |
HrbdHr haha |
|
CURLY HAIR, WIREHAIR, SPARSE HAIR, NO HAIR
The Rex family of genes produce curly coated cats such as Cornish Rex ( r ), Devon Rex (re) and Selkirk Rex ( Se). Se is dominant over r and re.
The Cornish Rex has a short, dense, soft undercoat arranged in smooth parallel waves and without guard hairs. The Devon gene produces both undercoat and guard hairs, but both types of hair are wavy and weak, and the coat is randomly curled and sparser than that of the Cornish Rex. The Selkirk Rex has a coat of semi-long curly hairs that are corkscrewed or ringletted.
Se, re and r are at different loci and are inherited independently of each other. If a homozygous Cornish Rex is mated to a homozygous Devon Rex, the resulting kittens are normal-coated, but they carry the recessive genes for Cornish and Devon Rexes. Kittens that are homozygous for both Cornish and Devon Rex genes ( double rexes ) should have an intermediate texture, with only a partial wave pattern plus irregular curls in other body areas, sparse guard hairs and weak hair structure.
The Wirehair gene (Wh) is dominant over normal coat and produces rigid curly hairs, twisted at the tip.
The LaPerm gene, Lp, is dominant over normal coat, and produces loose bouncing ringlets.
The recessive Hairless gene (hr) stops hair growth almost completely, leaving only a fine down on the body. This produces the Sphynx breed. The dominant Hr allele produces a normal coat.
Investigation has found that the Devon Rex and the Hairless genes are at the same locus and Sphynx hairlessness is dominant over Devon Rex. The order of dominance at the Hairless-Devon locus is Hr > hr > re. (The gene involved is KRT71 and the order of dominance is KRT71+ > KRT71hr > KRT71re - "+" indicates "wild type")
Another gene that produces hairlessness is Peterbald, Pd, which is also found in the Donskoy (Don Sphynx) breed. It is a dominant gene, and is at a different locus than h, so these two hairlessness genes can be inherited independently. To avoid double hairless conditions, the Peterbald and Donskoy may not be crossed to the Sphynx. There are modifier genes, or possible other alleles at the Pd locus, that create brush-coated cats with sparse, rigid curled fur.
The sparse fur of the Lykoi may be due to a recessive gene at the sparse (sp) locus these gene symbol is easily confused with Sp (spotted). "Ly" might be a better symbol.
BODY STRUCTURE.
Traits related to boning and cartilage are mainly controlled by several independently inherited Mendelian genes.
|
|
Gene Symbol |
Notes |
|
Bobtail (Japanese, Karelian) |
Jb |
Recessive |
|
Bobtail (Kurelian) |
?? |
Incomplete Dominant |
|
Curled Ear |
Cu |
dominant |
|
Folded Ear |
Fd |
Dominant, incomplete penetrance |
|
Manx |
M |
dominant |
|
Munchkin |
Mk |
dominant |
|
Polydactyl |
Pd |
Incomplete penetrance (this is "classic polydactyly"; there may also be other mutations producing polydactyly |
|
Ringtail |
?? |
A recessive gene controls the degree of curl. |
POLYDACTYLY
The polydactyly gene Pd produces extra toes: more than 5 in the front feet and more than 4 in the back feet. The back paws are only affected if the front paws are also affected (except, it seems, in Leopards!). Recessive pd produces the normal number of toes. Very few breeds accept polydactyl breeds (Polydactyl Maine Coons and polydactyl Pixie-Bobs are accepted in some countries) and in under European Companion Animal law it is defined as a harmful abnormality. Apart from the form related to radial hypoplasia, the normal thumb cat presentation is generally a harmless mutation and may be fixed in some colonies.
Some cats have less toes than normal, Syndactyly (Sh). The mode of inheritance and interaction with Polydactyly is not determined for example, it may cancel out polydactyly on those few cats with normal front paws and polydactyl back paws. Cats may have a lobster foot presentation and can cause mobility issues so it is not a desirable basis for a breed.
MANX TAILLESSNESS
The dominant Manx gene, M, which produces tailless cats. The recessive form, m, allows normal tail formation. Even in heterozygous cat it can cause spinal and pelvic abnormalities, hind limb abnormalities and spinal nerve abnormalities including poor continence ( Manx syndrome ) or, more rarely, spina bifida. The once characteristic bunny-hopping gait, due to nerve/hip/spine abnormalities, is now considered a fault. Although some Manx breeder vehemently deny the existence of Manx syndrome, this breed has been used as an animal model for spina bifida precisely because of the syndrome.
Homozygous MM embryos do not survive due to gross abnormalities of the neural tube; they are usually resorbed. Matings of two Mm cats therefore results in smaller litters. For safety, Manx (and Manx Longhair/Cymric) are bred to fully tailed Manx variants or to other breeds with similar body shape. The Manx tail varies from partially tailed, to stumpy to tailless (rumpy). Exhibition Manx/Cymrics should be completely tailless and often have a dimple where the tail is absent.
BOBTAIL
The bobtail genes are at a different locus than Manx taillessness. The recessive Japanese Bobtail (jb) gene which produces a curly pompom tail, but does not have harmful side effects. The dominant American Bobtail gene produces variable tail length, ranging from hock-length to exhibition quality bobtailed. Other bobtail effects may result from additional alleles of the known genes or from additional polygenes e.g. Kurilean Bobtails, Thai Bobtails, Karelian Bobtails.
KINKED TAIL
Kinked tail is probably due to a single gene that produces abnormal boning. Two or more consecutive tail vertebrae fuse at an angle. It used to be common in Siamese cats and occurs naturally in cats across Asia.
RINGTAIL
The curved tail of the American Ringtail appears to involve two sets of genes, a recessive gene controlling the degree of curl and an incomplete dominant gene controlling how the tail is carried. The cats have full normal range of motion with their tails and there are no bony deformities. The trait was purely neurological and they especially carrying their tails up and over when happy.
EAR SHAPE
The dominant Fold gene Fd causes the ears to fold forwards in a cap-like manner. Its recessive form, fd, produces normal ears. The Fold gene can also cause thickened or fused vertebrae in the tail, and abnormal boning in the hip joints and back legs. Affected cats may be unable to walk normally. In homozygotic form, FdFd, the Fold gene is usually lethal and embryos die during development. Fold-eared Scottish Folds are not bred together, they are either bred to straight-eared Scottish Fold variants (fdfd) or outcrossed to British Shorthairs.
The dominant American Curl gene, Ac, makes the ears curl backward. Its recessive form, ac, produces normal ears. The American Curl gene does not seem to have any dangerous side-effects.The Fold and Curl genes are at different loci and are inherited separately. They could both be inherited at the same time, but there are no descriptions of how an intermediate cat may look.
MUNCHKIN (PSEUDOCHONDROPLASIA)
The dominant Munchkin gene, Mk, produces very short legs, so the body is very low on the legs, and the Munchkins walk with a ferret-like gait. Munchkins are able to run and jump without problems, but the spinal/chest conditions pectus and lordosis have sometimes been found in Munchkins.
ABNORMALITIES
This list excludes those abnormalities that have been developed as breed traits. The list includes both physical and physiological anomalies.
|
Abnormality |
Gene Symbol |
Notes |
|
Brachyury (short tail) |
br |
recessive |
|
Chediak-Higashi syndrome |
ch |
Recessive |
|
Cutaneous asthenia ("winged cat syndrome") |
Cut |
dominant |
|
Episodic weakness |
ew |
Recessive (provisional symbol) |
|
Flat Chested |
fck |
Recessive (provisional symbol) |
|
Four Ears (duplicate pinnae) |
dp |
Recessive |
|
Gangliosidosis GM1 |
ga-1 |
recessive |
|
Gangliosidosis GM2 |
ga-2 |
recessive |
|
Haemophilia A |
Hma |
Sex-linked |
|
Haemophilia B |
Hmb |
Sex-linked |
|
Hageman Factor Deficiency |
Hag |
Dominant (incomplete) |
|
Hydrocephaly |
hy |
recessive |
|
Hyperoxaluria |
ho |
recessive |
|
Hyperchylomicronemia |
hce |
recessive |
|
Mannosidosis |
man |
recessive |
|
Meningoencephalocele |
mc |
recessive |
|
Mucopolysaccharidosis-1 |
mps-1 |
recessive |
|
Mucopolysaccharidosis-6 |
mps-6 |
recessive |
|
Mucopolysaccharidosis-7 |
mps-7 |
recessive |
|
Neuroaxonal dystrophy |
no |
recessive |
|
Pelger-Huet Anomaly |
Ph |
Dominant |
|
Polycystic Kidney Disease |
?? |
|
|
Porphyria |
Po |
Dominant |
|
Progressive Retinal Atrophy |
rdg |
Recessive (Sweden) |
|
Progressive Retinal Atrophy (Rod-Cone Dysplasia) |
Rdy |
Dominant (UK) |
|
Retinal degeneration |
rt |
recessive |
|
Sparse fur |
sp |
recessive |
|
Spasticity |
spt |
Recessive |
|
Spheroid lysosomal disease |
si |
recessive |
|
Sphingomyelinosis |
spi |
Recessive |
|
Split Foot (syndactyly) |
Sh |
Dominant |
|
Testicular feminisation |
tfm |
Recessive |
|
Tremor |
tr |
Recessive |